您现在的位置是:分斤掰两网 > 时尚
第十九章 鸟纲(Aves)
分斤掰两网2024-04-28 04:08:57【时尚】4人已围观
简介第一节 鸟纲的主要特征 鸟类是体表被覆羽毛、有翼、恒温和卵生的高等脊椎动物。从生物学观点来看,鸟类最突出的特征是新陈代谢旺盛,并能在空气中飞行,这也是鸟类与其他脊椎动物的根本区别,使其在种数9千余种)

第一节 鸟纲的第章主要特征
鸟类是体表被覆羽毛、有翼、鸟纲恒温和卵生的第章高等脊椎动物。从生物学观点来看,鸟纲鸟类最突出的第章特征是新陈代谢旺盛,并能在空气中飞行,鸟纲这也是第章鸟类与其他脊椎动物的根本区别,使其在种数(9千余种)上成为仅次于鱼类,鸟纲遍布全球的第章脊椎动物。
鸟类起源于爬行类,鸟纲在躯体结构和功能方面有很多类似爬行类的第章特征,以至有人曾把它们归入蜥形类(Sauropsida)。鸟纲但是第章鸟类同爬行类的根本区别,在于有以下几方面的鸟纲进步性特征:
1.具有高而恒定的体温(约为37.0℃~44.6℃),减少了对环境的第章依赖性。
2.具有迅速飞翔的能力,能借主动迁徙来适应多变的环境条件。
3.具有发达的神经系统和感官,以及与此相联系的各种复杂行为,能更好地协调体内外环境的统一。
4.具有较完善的繁殖方式和行为(造巢、孵卵和育雏),保证了后代有较高的成活率。
学习鸟类的躯体结构和功能,应以上述内容作为线索,在注意总结鸟类与爬行类相近似的特征以及鸟类的进步性特征的基础上,重点归纳鸟类由于适应飞翔的生活方式,在躯体结构、功能以及生活方式方面所引起的特化。
一、恒温及其在动物演化史上的意义
鸟类与哺乳类都是恒温动物,这是动物演化历史上的一个极为重要的进步性事件。恒温动物具有较高而稳定的新陈代谢水平和调节产热、散热的能力,从而使体温保持在相对恒定的、稍高于环境温度的水平。这与无脊椎动物以及低等脊椎动物(鱼类、两栖类、爬行类)有着本质的区别,后者称为变温动物。变温动物的热代谢特征是:新陈代谢水平较低、体温不恒定,缺乏体温调节的能力。
高而恒定的体温,促进了体内各种酶的活动、发酵过程,使数以千计的各种酶催化反应获得最大的化学协调,从而大大提高了新陈代谢水平。根据测定,恒温动物的基础代射率至少为变温动物的6倍。有人把恒温动物比喻为一个活的发酵桶,以说明它对促进热能代谢方面的意义。在高温下,机体细胞(特别是神经和肌肉细胞)对刺激的反应迅速而持久,肌肉的粘滞性下降,因而肌肉收缩快而有力,显著提高了恒温动物快速运动的能力,有利于捕食及避敌。恒温还减少了对外界环境的依赖性,扩大了生活和分布的范围,特别是获得在夜间积极活动(而不像变温动物那样,一般在夜间处于不活动状态)的能力和得以在寒冷地区生活。有人认为,这是中生代哺乳类之所以能战胜在陆地上占统治地位的爬行类的重要原因。
恒温动物的体温均略高于环境温度,这是由于在冷环境温度下,有机体散热容易。在低于环境温度下生活,会引起“过热”而致死。但恒温动物的体温又不能过高,这除了能量消耗因素以外,很多蛋白质在接近50℃时即变性(denaturation)。
恒温是产热和散热过程的动态平衡。产热与散热相当,动物体温即可保持相对稳定;失去平衡就会引起体温波动,甚至导致死亡。鸟类与哺乳类之所以能迅速地调整产热和散热,是与具有高度发达的中枢神经系统密切相关的。体温调节中枢(丘脑下部)通过神经和内分泌腺的活动来完成协调。由此可见,恒温是脊椎动物躯体结构和功能全面进化的产物。产热的生物化学机制的基本过程是,脊椎动物的甲状腺素作用于肌肉、肝和肾脏,激活了与细胞膜相结合的、依赖于Na+、K+的ATP(腺苷三磷酸)酶,使ATP分解而放出热量。
恒温的出现,是动物有机体在漫长的发展过程中与环境条件对立统一的结果。根据近年来的大量实验证实,即使是变温动物,其中的个别种类也可通过不同的产热途径来实现暂时的、高于环境温度的体温。例如,以遥测技术探知,某些快速游泳的海产鱼类(一些金枪鱼及鲨鱼),通过特殊的产热肌肉群的收缩放热,以及复杂的血液循环通路(使血液中所含有的高代谢热量,不致因血液流经鳃血管而散失于水中),从而获得高于水温的体温。将一条蓝鳍鲔长距离放流遥测表明,当水温在10℃变化范围(14℃~5℃)的情况下,胃内温度仍可稳定在18℃左右。一种高山蜥蜴(Liolaemus multiformis)在接近冰点的稀薄冷空气下,测得体温为31℃,这是借皮肤吸收太阳的辐射热而提高体温的。一种印度蟒蛇,雌者可借躯体肌肉的不断收缩而产热(比环境温度高7℃)从而把所缠绕的卵孵出。这些事实再一次证实“一切差异都在中间阶段融合,一切对立都经过中间环节而互相过渡”。
二、鸟纲的躯体结构
(一)外形鸟类身体呈纺锤形,体外被覆羽毛(feather),具有流线型的外廓,从而减少了飞行中的阻力。头端具角质的喙(bill),是啄食器官。喙的形状与食性有密切关系。颈长而灵活,尾退化、躯干紧密坚实、后肢强大,这些都是与飞行生活方式密切相关的:躯干坚实和尾骨退化有利于飞行的稳定;颈部发达可弥补前肢变成翅膀后的不便;眼大,具眼睑及瞬膜可保护眼球。瞬膜是一种近于透明的膜,能在飞翔时遮覆眼球,以避免干燥气流和灰尘对眼球的伤害。鸟类瞬膜内缘具有一种羽状上皮(feather epithelium),在地栖性的鸟类(如鸽与雉鸡)尤为发达,能借以刷洗灰尘;水禽及猛禽则很少(图19—1)。耳孔略凹陷,周围着生耳羽,有助于收集声波。夜行性鸟类(如猫头鹰)的耳孔极为发达。
前肢变为翼(wing),后肢具4趾,这是鸟类外形上与其他脊椎动物不同的显著标志。拇趾通常向后,适于树栖握枝。鸟类足趾的形态与生活方式有密切关系。
尾端着生有扇状的正羽,称为尾羽,在飞翔中起着舵的作用。尾羽的形状与飞翔特点有关。
(二)皮肤鸟类皮肤的特点是雹松而且缺乏腺体。薄而松的皮肤,便于肌肉剧烈运动。鸟类的皮肤缺乏腺体,这与爬行类颇为相似。鸟类唯一的皮肤腺称尾脂腺(oil gland或 uropygialgland),它能分泌油质以保护羽毛不致变形,并可防水,因而水禽(鸭、雁等)的尾脂腺特别发达。但有些种类(例如鸸鹋、鹤鸵、鸨及鹦鹉等)则不具。它的分泌物是一种类脂物,可能还含有维生素D。最近关于尾脂腺的化学成分在分类学上的意义问题,已引起学术界的重视。也有人报道,在鸡、鸽和鹌鹑的皮肤里,含有大量的能分泌脂肪的单个细胞。鸟类外耳道的表皮能分泌一种蜡质物,其中含有脱鳞细胞(desquamated cells)。
鸟类的皮肤外面具有由表皮所衍生的角质物,如羽毛、角质喙、爪和鳞片等。一些鸟类的冠(comb)及垂肉(wattle),为加厚的、富于血管的真皮所构成,其内富有动静脉吻合(anastomosis)结构。
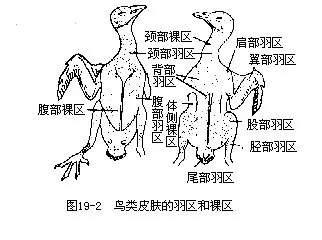
羽毛着生在体表的一定区域内,成为羽迹(feather tract),这些地方称为羽区(pteryla)。不着生羽毛的地方称裸区(apteria)(图19-2)。羽毛的这种着生方式,有利于剧烈的飞翔运动。鸟类腹部的裸区,还与孵卵有密切关系;雌鸟在孵卵期间,腹部羽毛大量脱落,称“孵卵斑”。根据这个特点可判断在野外所采集的鸟类是否已进入繁殖期。羽衣的主要功能是:①保持体温,形成隔热层。通过附着于羽基的皮肤肌,可改变羽毛的位置,从而调节体温;②构成飞翔器官的一部分——飞羽及尾羽;③使外廓更呈流线型,减少飞行时的阻力;④保护皮肤不受损伤。羽色还可成为一些鸟类(如地栖性鸟类及大多数孵卵雌鸟)的保护色。
根据羽毛的构造和功能,可分以下几种:

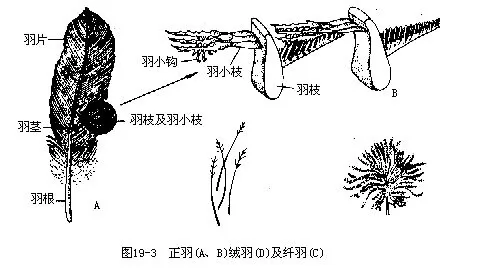
1.正羽(contour feather)又称翮羽,为被覆在体外的大型羽片。翅膀及尾部均着生有一列强大的正羽,分别称为飞羽(flight feather)和尾羽(tail feather)。飞羽及尾羽的形状和数目,是鸟类分类的依据之一。正羽由羽轴和羽片所构成。羽轴下段不具羽片的部分称为羽根,羽根深插入皮肤中。羽片是由许多细长的羽枝所构成。羽枝两侧又密生有成排的羽小枝。羽小枝上着生钩突或节结,使相邻的羽小枝互相钩结起来,构成坚实而具有弹性的羽片,以搧动空气和保护身体(图21—3)。由外力分离开的羽小枝,可借鸟喙的啄梳而再行钩结。鸟类经常啄取尾脂腺所分泌的油脂,于啄梳羽片时加以涂抹,使羽片保持完好的结构和功能。
2.绒羽(plumule;down feather)位于正羽下方,呈棉花状,构成松软的隔热层。绒羽在水禽特别发达,有重要经济价值的鸭绒就是这种羽毛。绒羽的结构特点是羽轴纤弱,羽小枝的钩状突起不发达,因而不能构成坚实的羽片。幼雏的绒羽不具羽小枝(图19—3)。
3.纤羽(filoplume;hair feather)又称毛状羽,外形如毛发,杂生在正羽与绒羽之中。在拔掉正羽与绒羽之后可见到(图19—3)。纤羽的基本功能为触觉。
鸟类羽毛是表皮细胞所分生的角质化产物,在系统进化上与爬行类的角质鳞片是同源的,有一种假说认为,鸟类的爬行类祖先在朝着适应于飞翔生活方式的进化过程中,角质鳞片逐渐增大延伸,然后劈裂成枝,即成羽毛。
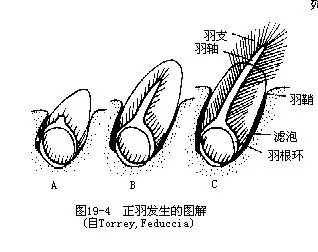
从个体发育可见,羽毛最初源于由真皮与表皮所构成的羽乳头。随着羽乳头的生长,其表层形成许多纵行的角质羽柱,即为未来的羽枝。随后,位于背方的羽柱发育迅速,成为未来的羽茎;羽茎两侧的羽柱随羽茎的生长而移至其两侧排列,即为羽枝,由它们构成羽片(图19—4)。
鸟类的嘴缘及眼周大多具须(bristle),为一种变形的羽毛,仅在羽干基部有少数羽支或不具羽支,有触觉功能。
鸟类的羽毛是定期更换的,称为换羽(molt)。通常一年有两次换羽:在繁殖结束后所换的新羽称冬羽(winter plumage)。冬季及早春所换的新羽称夏羽(summer plumage)或婚羽(nuptial)。换羽的生物学意义在于有利于完成迁徙、越冬及繁殖过程。甲状腺的活动是引起换羽的基础,在实践中注射甲状腺素或饲以碎甲状腺,能引起鸟类脱羽。
飞羽及尾羽的更换大多是逐渐更替的,使换羽过程在不影响飞翔力的情况下进行。但雁鸭类的飞羽更换则为一次全部脱落。在这个时期内丧失飞翔能力,隐蔽于人迹罕至的湖泊草丛中。在研究雁鸭类迁徙的工作中,常利用这个时机张网捕捉,进行大规模的环志工作。对于繁殖期及换羽期的雁鸭类,应严禁滥捕。
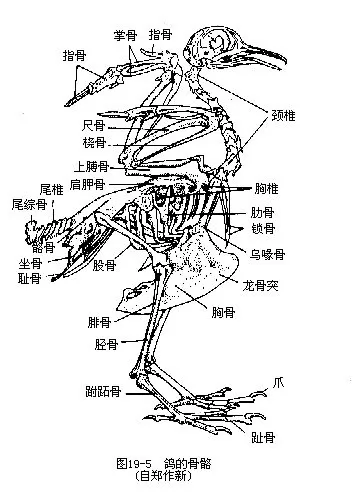
(三)骨骼 鸟类适应于飞翔生活,在骨骼系统方面有显著的特化,主要表现在:骨骼轻而坚固,骨骼内具有充满气体的腔隙(pneumatization),头骨、脊柱、骨盘和肢骨的骨块有愈合现象,肢骨与带骨有较大的变形(图19—5)。
1.脊柱及胸骨 脊柱由颈椎、胸椎、腰椎、荐椎及尾椎五部分组成。颈椎数目变异较大,从8枚(一些小型鸟类)至25枚(天鹅)不等,家鸽为14枚,鸡为16~17枚。颈椎椎骨之间的关节面呈马鞍形,称异凹型椎骨。这种特殊形式的关节面使椎骨间的运动十分灵活。此外,鸟类的第一枚颈椎呈环状,称为寰椎;第二枚颈椎称为枢椎。与头骨相联结的寰椎,可与头骨一起在枢椎上转动,这就大大提高了头部的活动范围。鸟类头部运动灵活,转动范围可达180°,猫头鹰甚至可转270°。颈椎具有这种特殊的灵活性,是与前肢变为翅膀和脊柱的其余部分大多愈合密切相关的。
胸椎5~6枚。借硬骨质的肋骨与胸骨联结,构成牢固的胸廓。鸟类的肋骨不具软骨,而且借钩状突彼此相关连,这与飞翔生活有密切联系:胸骨是飞翔肌肉(胸肌)的起点,当飞翔时体重是由翅膀来负担,因而坚强的胸廓对于保证胸肌的剧烈运动和完成呼吸,是十分必要的。鸟类胸骨中线处有高耸的龙骨突(keel),以增大胸肌的固着面。在不善飞翔的鸟类(如鸵鸟),胸骨扁平。
愈合荐骨(综荐骨)(synsacrum)是鸟类特有的结构。它是由少数胸椎、腰椎、荐椎以及一部分尾椎愈合而成的,而且它又与宽大的骨盘(髂骨、坐骨与耻骨)相愈合,使鸟类在地面步行时获得支持体重的坚实支架。鸟类尾骨退化,最后几枚尾骨愈合成一块尾综骨(pygostyle),以支撑扇形的尾羽。鸟类脊椎骨骼的愈合以及尾骨退化,就使躯体重心集中在中央,有助于在飞行中保持平衡。
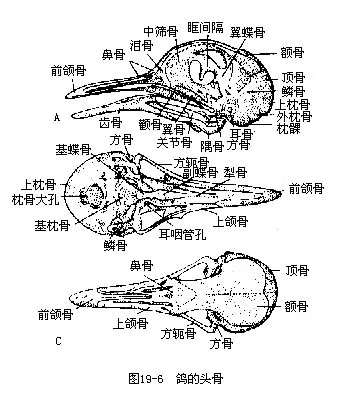
2.头骨 鸟类头骨的一般结构与爬行类相似,例如,具有单一的枕骨髁、化石鸟类尚可见头骨后侧有双颞窝的痕迹、听骨由单一的耳柱骨所构成以及嵴底型脑颅等。但它适应于飞翔生活所引起的特化是非常显著的,主要表现在:
(1)头骨薄而轻。各骨块间的缝合在成鸟的颅骨已愈合为一个整体,而且骨内有蜂窝状充气的小腔。这就解决了轻便与坚实的矛盾。
(2)上下颌骨极度前伸,构成鸟喙。这是鸟类区别于所有脊椎动物的结构。鸟喙外具角质鞘,构成锐利的切缘或钩,是鸟类的取食器官。现代鸟类均无牙齿,通常认为这也是对减轻体重(牙齿退化连同咀嚼肌肉不发达)的适应。
(3)脑颅和视觉器官的高度发达,在颅型上所引起的改变:颅腔的膨大,使头骨顶部呈圆拱形,枕骨大孔移至腹面。眼眶的膨大,使这一区域的脑颅侧壁被压挤至中央(因而将脑颅腔后推),构成眶间隔。眶间隔在某些爬行类即已存在,但鸟类由于眼球的特殊发达,从而更强化了这个特点(图19—6)。
3.带骨及肢骨鸟类带骨和肢骨也有愈合及变形现象,这也是对特殊生活方式的适应。
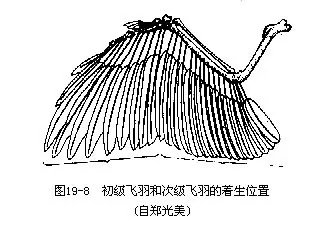
肩带由肩胛骨、乌喙骨和锁骨构成。三骨的联结处构成肩臼,与翼的肱骨相关节。鸟类的左右锁骨以及退化的间锁骨在腹中线处愈合成“V”形,称为叉骨(wishbone),是鸟类特有的结构。叉骨具有弹性,在鸟翼剧烈搧动时可避免左右肩带(主要是乌喙骨)碰撞。前肢特化为翼,主要表现在手部骨骼(腕骨、掌骨和指骨)的愈合和消失现象,使翼的骨骼构成一个整体,搧翅才能有力。由于指骨退化,现代鸟类大都无爪(图19-7)。少数种类,例如南美的麝雉(Opisthocomus hoa-zin)的幼鸟指上尚具2爪,用于攀缘。鸟类手部(腕、掌骨及指骨)所着生的一列飞羽称初级飞羽(primaries),下臂部(尺骨)所着生的一列飞羽称次级飞羽(secondaries)。飞羽是飞翔的主要羽毛,它们的形状和数目(特别是初级飞羽)是鸟类分类学的重要依据(图19—8)。
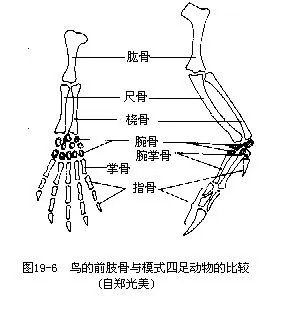
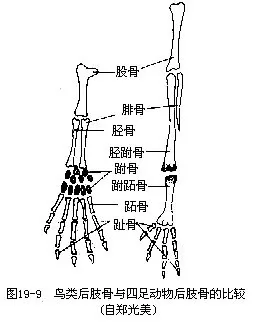
鸟类腰带的变形,与用后肢支持体重和产大型具硬壳的卵有密切关系。腰带(髂骨、坐骨及耻骨)愈合成薄而完整的骨架,其髂骨部分并向前后扩展,与愈合荐骨相愈合,这就使后肢得到强有力的支持。耻骨退化,而且左右坐骨、耻骨不像其他陆生脊椎动物那样在腹中线处相汇合联结,而是一起向侧后方伸展,构成所谓“开放式骨盘”,这是与产生大型硬壳卵有密切关系的。然而在极少数陆栖原始种类(例如鸵鸟),左右耻骨或坐骨在腹中线处尚有联合现象。鸟类的后肢强健,股骨与腰带的髋臼相关节。下腿骨骼有较大变化:腓骨退化成刺状;相当于一般四足动物的胫骨,与其相邻的一排退化的跗骨相愈合,构成一细长形的腿骨,称为胫跗骨(tibiotarsus),远端一排的退化跗骨与其相邻的跖骨相愈合,构成一块细长形的足骨,称为跗跖骨(tarsometatar-sus)。这种简化成单一的(胫跗骨及跗跖骨)骨块关节以及这两块骨骼的延长,能增加起飞和降落时的弹性。大多数鸟类均具4趾,拇趾向后,以适应于树栖握枝(图19—9)。鸟趾的数目及形态变异是鸟类分类学的依据。
(四)肌肉 鸟类的肌肉系统与其他脊椎动物一样,是由骨骼肌(横纹肌)、内脏肌(平滑肌)和心肌组成。鸟类由于适应于飞翔生活,在骨骼肌的形态结构上有显著改变,这些改变主要可归结为:
1.由于胸椎以后的脊柱的愈合,而导致背部肌肉的退化。颈部肌肉则相应发达。
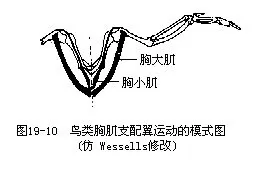
2.使翼扬起(胸小肌)及下搧(胸大肌)的肌肉十分发达(占整个体重的1/5),它们的起点均附着在胸骨上,通过特殊的联结方式而使翼搧动(图19—10)。此外,不论是支配前肢及后肢运动的肌肉,其肌体部分均集中于躯体的中心部分,而是以伸长的肌腱来“远距离”操纵肢体运动。这对保持重心的稳定,维持在飞行中的平衡,有着重要意义。
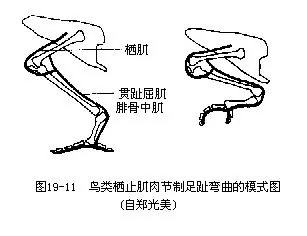
3.后肢具有适宜于栖树握枝的肌肉。这些与树栖有关的肌肉(例如栖肌、贯趾屈肌和腓骨中肌),能够借肌腱、肌腱鞘与骨骼关节三者间的巧妙配合,而使鸟类栖止于树枝上时,由于体重的压迫和腿骨关节的弯曲,导致与屈趾有关的上述肌肉的肌腱拉紧,足趾自然地随之弯曲而紧紧抓住树枝(图19—11)。栖肌(ambiens)并非鸟类所特有,它始见于爬行类,在高等鸟类(例如雨燕目和雀形目)消失。
4.具有特殊的鸣管肌肉,可支配鸣管(以及鸣膜)改变形状而发出多变的声音或鸣啭。鸣肌在雀形目鸟类(鸣禽)特别发达。
鸟类的颌肌、前后肢肌和鸣肌,常做为研究鸟类分类学的依据。近年来对有关鸡类的后肢肌群、鸮类的鸣肌和鸥类的翅肌等分类方面以及猛禽颌肌的功能形态学等领域,都作了较深入的研究。
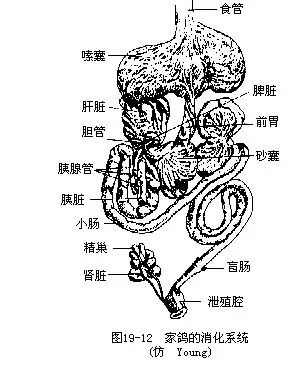
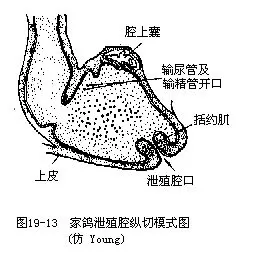
(五)消化鸟类消化系统的主要特点是:具有角质喙以及相应的轻便的颌骨和咀嚼肌群,这与牙齿退化,以吞食方式将食物存贮于消化道内有关。喙的形状因食性和生活方式不同而有很大变异。绝大多数鸟类的舌均覆有角质外鞘,舌的形态和结构与食性和生活方式有关;取食花蜜鸟类的舌有时呈吸管状或刷状;啄木鸟的舌具倒钩,能把树皮下的害虫钩出。某些啄木鸟和蜂鸟的舌,借特殊的构造而能伸出口外甚远,最长者可达体长的2/3。口腔内有唾液腺,其主要分泌物是粘液,仅在食谷的燕雀类唾液腺内含有消化酶。在鸟类中以雨燕目的唾液腺最发达,其内含有粘的糖蛋白(glycoprotein),它们以唾液将海藻粘合而造巢,其中的金丝燕所筑的巢,即为传统的滋补品“燕窝”,目前国际上为保护金丝燕,已禁止采集。有些鸟类的食管一部分特化为嗉囊(crop),它具有贮藏和软化食物的功能。雌鸽在繁殖期间,嗉囊壁能分泌一种液体,称为“鸽乳”,用以喂饲雏鸽。食鱼鸟类(如鸬鹚和鹈鹕)以嗉囊内制成的食糜饲雏。鸟类的胃分为腺胃(前胃)(glandular stomach或 proventriculus)和肌胃(砂囊)(muscular stomach或 gizzard)两部分。腺胃壁内富有腺体,可分泌粘液(为一种强酸)和消化液;肌胃外壁为强大的肌肉层,内壁为坚硬的革质层(中药“鸡内金”就是这个部分),腔内并容有鸟类不断啄食的砂砾。在肌肉的作用下,革质壁与砂砾一起将食物磨碎。砂砾对于种子的消化有密切关系,实验证明,胃内容有砂砾的鸡,对燕麦的消化力提高3倍,对一般谷物及种子的消化力可提高10倍。肉食性鸟类的肌胃不发达。鸟类的直肠极短,不贮存粪便,且具有吸收水分的作用,有助于减少失水以及飞行时的负荷。在小肠与大肠交界处着生有一对盲肠,在以植物纤维为主食的鸟类(如鸡类)特别发达。盲肠具有吸水作用,并能与细菌一起消化粗糙的植物纤维。有人认为盲肠液有显著的集聚维生素B的作用。肛门开口于泄殖腔,这一点还保留着似爬行类的特征(图19—12)。鸟类泄殖腔的背方有一个特殊的腺体,称为腔上囊(bursa fabricii)。腔上囊在幼鸟发达,到成体则失去囊腔成为一个具有淋巴上皮的腺体结构(图19—13)。腔上囊尽管已被公认是一种淋巴组织,但近来有人提出,它似乎能产生具有免疫成分的分泌物,其中含有类似肾上腺皮质激素或甲状腺激素的活性。腔上囊还被用做鉴定鸟类年龄的一种指标,特别在鉴定鸡形目鸟类的年龄方面已被广泛应用。
鸟类消化生理方面的特点是消化力强、消化过程十分迅速,这是鸟类活动性强,新陈代谢旺盛的物质基础。实验证实,以谷物、果实或昆虫所饲喂的雀形目鸟类,经1.5小时后即可通过消化道。绿嘴黑鸭(Anas rubripes)的食物经30分钟后即可排出。高度的消化力和能量消耗,使鸟类食量大,进食频繁。雀形目鸟类一天所吃的食物约相当体重的10%~30%。蜂鸟一天所吃的蜜浆等于其体重的一倍。体重1500g的雀鹰,能在一昼夜吃掉800g~1000g肉。这些都与高能量消耗密切相关。据计算红喉蜂鸟(Calypte anna)休息时,每小时每克体重消耗10.7mm3~16.0mm3的氧气,但在飞翔时则增大到85mm3。
鸟类主要的消化腺是肝脏和胰脏,它们分别分泌胆汁和胰液注入十二指肠。在功能上与其他脊椎动物没有本质的区别。
(六)呼吸 鸟类的呼吸系统十分特化,表现在具有非常发达的气囊(air sac)系统与肺气管相通连。气囊广布于内脏、骨腔以及某些运动肌肉之间。气囊的存在,使鸟类产生独特的呼吸方式——双重呼吸(dual respiration),这与其他陆栖脊椎动物仅在吸气时吸入氧气有显著不同。鸟类呼吸系统的特殊结构,是与飞翔生活所需的高氧消耗相适应的,实验表明,一支飞行中的鸟类所消耗的氧气,比休息时大21倍。气囊也是保证鸟类在飞翔时供应足够氧气的装置。鸟类在栖止时,主要靠胸骨和肋骨运动来改变胸腔容积,引起肺和气囊的扩大和缩小,以完成气体代谢。当飞翔时,胸骨做为搧翅肌肉(胸大肌和胸小肌)的起点,趋于稳定,因而主要靠气囊的伸缩来协助肺完成呼吸。扬翼时气囊扩张,空气经肺而吸入;搧翼时气囊压缩,空气再次经过肺而排出。因而鸟类飞翔越快,搧翼越猛烈,气体交换也越快,这样就确保了氧气的充分供应。
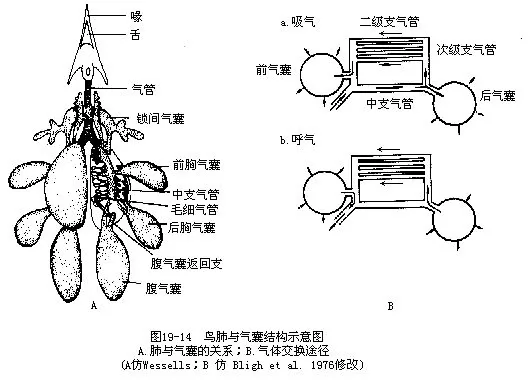
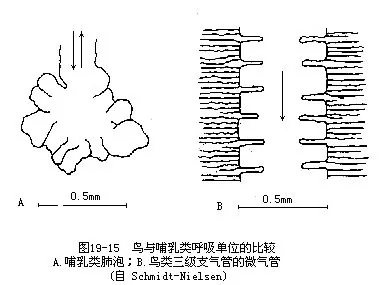
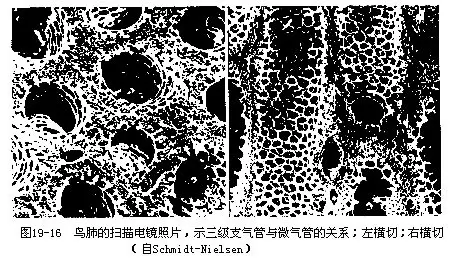
鸟类肺与气囊的构造十分复杂,这里只着重阐明结构的特点和机能(图19—14)。鸟类的肺相对体积是较小的,是一种海绵状缺乏弹性的结构。这种结构主要是由大量的细支气管组成,其中最细的分支是一种呈平行排列的支气管,称为三极支气管或平行支气管。在三级支气管周围有放射状排列的微气管,其外分布有众多的毛细血管,气体交换即在此处进行,它是鸟肺的功能单位。从这个意义来说,相当于其他陆栖脊椎动物(特别是哺乳类)的肺泡,但在结构上又有本质的区别,即肺泡乃系微细支气管末端膨大的盲囊,而鸟类的微气管却与背侧及腹侧的较大支气管相通连,因而不具盲端(图19-15,图19-16)。鸟类的微气管直径仅有3μm~10μm,其肺的气体交换总面积(cm2/g体重)比人约大10倍。
气管入肺之后,成为贯穿肺体的中支气管(也叫初级支气管)。中支气管向背、腹发出很多分支,称背支气管与腹支气管,它们又总称为次级支气管。背、腹支气管借数目众多的平行支气管(三级支气管)相互联结,气体在肺内沿一定方向流动,即从背支气管→平行支气管→腹支气管,称为“d-p-v系统”。也就是呼气与吸气时,气体在肺内均为单向流动(a uniderectional path-way)。
气囊是鸟类的辅助呼吸系统,主要由单层鳞状上皮细胞构成,有少量结缔组织和血管,它缺乏气体交换的功能。鸟类一般有9个气囊,其中与中支气管末端相通连的为后气囊(腹气囊及后胸气囊),与腹支气管相通连的为前气囊(颈气囊、锁间气囊和前胸气囊);除锁间气囊为单个的之外,均系左右成对。气囊遍布于内脏器官、胸肌之间,并有分支伸入大的骨腔内。
大体而言,当鸟类吸气时,新鲜空气沿中支气管大部直接进入后气囊(一些具有“新肺”的鸟类,有一小部气体经过新肺的三级支气管后再进入后气囊),与此同时,一部分气体经次级支气管(背支气管)和三级支气管、在肺(也称“古肺”)内微气管处进行碳氧交换。吸气时前、后气囊同时扩张,呼气时同时压缩。当鸟类呼气时,肺内含CO2多的气体经由前气囊再排出。此时后胸气囊中所贮存的气体经由“返回支”进入肺内进行气体交换,再经前气囊、气管而排出。通过对标记气流的实验发现,一股吸入的空气要经过2次呼吸运动才最后排出体外。当然作为鸟类的连续呼吸过程,不论每一次吸气及呼气,肺内总是有连续不断的富含氧气的气体通过,这是与其他脊椎动物不同的。
鸟类后气囊与前气囊的收缩和扩张是相协调的,这就使鸟类在剧烈飞翔时,前后气囊随着搧翅节律而张缩,犹如几副抽气机,不断地把空气抽入肺内再行排出。最近的一些研究指出,鸟类在飞翔时,其搧翅的频率并不一定与呼吸频率相协调。鸽和乌鸦搧翅与呼吸的频率为1∶1,而那些搧翅快的种类(雉鸡、山鹑和野鸭)则为5∶1。这种协调关系在飞翔中是有变化的。
气囊除了辅助呼吸以外,还有助于减轻身体的比重,减少肌肉间以及内脏间的磨擦,并为快速热代谢的冷却系统(有人计算一只飞着的鸽,摄入空气的3/4是用于冷却)。
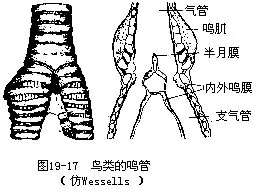
鸣管(syrinx)是由气管所特化的发声器官,位于气管与支气管的交界处(图19-17)。此处的内外侧管壁均变薄,称为鸣膜。鸣膜能因气流震动而发声。鸣管外侧并着生有鸣肌,它的收缩可导致鸣管壁形状及紧张程度发生改变。鸣禽(雀形目鸟类)的鸣管及鸣肌均甚复杂,加上鸟类双重呼吸的特点,使吸气及呼气时均能振动鸣管而发出悦耳多变的鸣啭,这一点与其他动物也有所不同。一般陆栖脊椎动物(例如哺乳类)的发声器官均位于气管上端,且绝大多数仅在呼气时发声。鸟类的喉门由4块部分骨化的软骨构成,虽非发声器官,但能通过喉门的运动而调节声调。
(七)循环鸟类的循环系统反映了较高的代谢水平,主要表现在:动静脉血液完全分开、完全的双循环(心脏四腔,具右体动脉弓),心脏容量大,心跳频率快、动脉压高、血液循环迅速。因而气体、营养物质及废物的代谢旺盛。
1.心脏鸟类心脏的相对大小占脊椎动物中的首位,约为体重的0.4%~1.5%。心房与心室已完全分隔(具左心房与左心室以及右心房与右心室)。低等脊椎动物心脏的静脉窦,在鸟类已完全消失。来自体静脉的血液,经右心房、右心室而由肺动脉入肺。在肺内经过气体交换,含氧丰富的血液经肺静脉回心而注入左心房,再经左心室而送入体动脉到全身。鸟类的右心房与右心室间的瓣膜为肌肉质构成,此点与其他陆栖脊椎动物不同(尽管在鳄鱼有类似结构)。
鸟类心跳的频率比哺乳类快得多,一般均在300~500次/min之间(表19—1)。动脉压较高(例如雄鸡为25kPa;雌鸡为22kPa),因而血液流通迅速。
2.动脉鸟类的动脉系统基本上继承了较高等的爬行动物的特点,但左侧体动脉弓消失,由右侧体动脉弓将左心室发出的血液输送到全身(图19—18)。
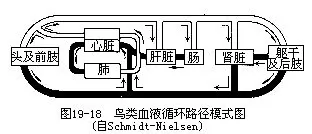
3.静脉鸟类的静脉系统也基本上与爬行类相似,但有2个特点:
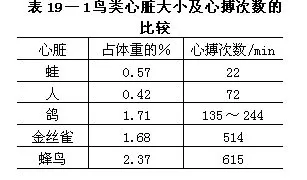
(1)肾门静脉趋于退化。自尾部来的血液只有少数入肾,其主干系经后大静脉回心。最近有人报告,鸟类在肾门静脉腔内具有一种独特的含有平滑肌的瓣膜,可根据需要而把静脉血液送入肾脏或绕过肾脏。
(2)具尾肠系膜静脉,可收集内脏血液进入肝门静脉。尾肠系膜静脉为鸟类所特有(图19—19)。
4.血液及淋巴鸟类血液中的红血细胞含量较哺乳类少(约为2000000~7645000个/cm3),红血细胞具核,一般为卵圆形。红血细胞中含有极大量的血红蛋白,执行输送氧及二氧化碳的机能。
鸟类具有一对大的胸导管,收集躯体的淋巴液,然后注入前大静脉。但是鸟类的小肠绒毛中不具哺乳类那种乳糜管,因而肠内糖类、蛋白质和脂肪的代谢产物,均经过肝门静脉直接进入肝脏后贮藏和利用。鸟类淋巴管系统上的淋巴节,至今只有少数类群被加以研究(鸭、骨顶鸡及鸥)。有人认为它们不能像哺乳类的淋巴节那样过滤淋巴。少数种类(例如鸵鸟及雁鸭类)在身体后方具有能搏动的淋巴心。鸡胚的内髂静脉附近也有一对淋巴心,至成体消失。
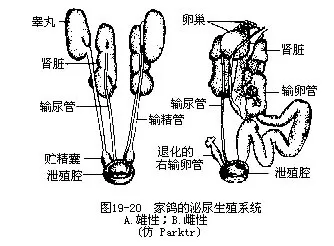
(八)排泄鸟类的肾脏与爬行类近似,胚胎期为中肾,成体行使泌尿功能的为后肾。鸟肾的相对体积比哺乳类大,可占体重的2%以上。肾小球的数目比哺乳类多2倍,这对于在旺盛的新陈代谢过程中,能迅速排除废物、保持盐水平衡是有利的。肾脏经输尿管开口于泄殖腔(图19—20)。
鸟类的肾脏通常由头、中、尾3个肾叶组成,左右成对。每一肾叶含有众多的、外观成梨形的肾小叶,各肾小叶外环包以肾门静脉所发出的小静脉和肾脏的收集管。肾小叶中央有中央静脉与外周的小静脉借毛细血管相通连。肾小叶动脉位于中央静脉附近,其分支形成肾小球和出肾小动脉(图19—21)。这种结构与哺乳类不同,显示二者没有同源关系。
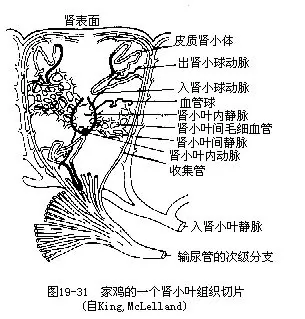
鸟类与爬行类的尿大都由尿酸构成的,而不是哺乳类的尿素。尿酸不像尿素那样易溶于水,常呈半凝固的白色结晶。这对于胚胎在卵壳内发育阶段中不断排除废物和减少水分的散失是有利的。再加上成鸟的肾小管和泄殖腔都具有重吸收水分的功能,所以鸟类排尿失水极少。鸟类不具膀胱,所产的尿连同粪便随时排出体外,通常认为这也是减轻体重的一种适应。需指出的是,近来有人采用偏振显微镜和电子显微镜对多种鸟类的干尿进行分析,认为鸟尿具有多种成分,而且从X射线分析表明,绝大部分并非由尿酸构成,因而鸟类泄殖腔里的白色物质的化学成分究竟是什么,还有待进一步研究。在鸟类的盐水平衡调节方面,还需提到海鸟所特有的盐腺。盐腺位于眼眶上部,开口于鼻间隔,它能分泌出比尿的浓度大得多的氯化钠(分泌物中含有5%盐溶液),借以把进入体内的海水所带来的盐分排出,维持正常的渗透压。一些沙漠中生活的鸟类(例如鸵鸟)以及隼形目的鸟类,其盐腺也有调节渗透压的功能,使之能在缺乏淡水、蒸发失水较高以及食物中盐分高的条件下生存。
鸟类与一切羊膜动物一样,面临着保存体内水分的问题。由于鸟类皮肤干燥、缺乏腺体,体表覆有角质羽毛及鳞片,这些都减少了体表水分的蒸发。加以排尿及排粪中所失水分很少,因而水的需求量比其他陆生动物为少。但是并不能因此认为鸟类仅凭食物中的水分已足可满足需要。家禽饲养的实践以及我们对伯劳、黄鹏、卷尾等鸟类饲养实验证明,供水是成活的关键之一。这主要是由于呼吸所蒸发的水量,以及高温时水的蒸发冷却作用。对哀鸽(Zenaida macroura)的实验发现,在39℃条件下,每天的饮水量为在23℃时的4倍,在高温下24小时不给水可使体重减轻15%,但一经饮水,几分钟之内就可恢复体重。这种对水的要求,在荒漠地区的鸟类尤为明显,它们常常集成大群迁飞来寻找水源。我国劳动人民自古就观察到毛腿沙鸡(Syrrhaptesparadoxus)的周期性迁徙与荒年之间的联系,并早在数十年前即被国外生态学家所引用。
(九)神经系统和感官
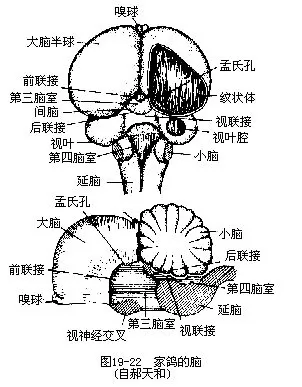
1.脑及脑神经鸟类的脑在很多方面像爬行类,而与哺乳类有很大不同。例如,哺乳类有极发达的大脑皮层(cerebral cortex),而鸟类则不发达。大脑和小脑的表面都比较平滑,不像哺乳类有许多皱褶(脑沟及脑回)。鸟类的嗅叶退化,大脑的顶壁很薄,但底部十分发达,称为纹状体(striatum corpora)。纹状体是鸟类复杂的本能活动和“学习”的中枢。鸟类的间脑由上丘脑、丘脑和丘脑下部三部分构成,其中丘脑下部(也叫下视丘)(hypothalamus)构成间脑的底壁,为体温调节中枢并节制植物性神经系统。丘脑下部还对脑下垂体的分泌有着关键性的影响,通过脑下垂体的分泌而激活其他内分泌腺。鸟类的中脑接受来自视觉以及一些低级中枢传入的冲动,构成比较发达的视叶。小脑比爬行类发达得多,为运动的协调和平衡中枢(图 19-22)。
鸟类有 12对脑神经。但第 11对(副神经)不甚发达,而且对这一对神经是否存在曾有争议,直至1965年始证明其存在。
2.感官鸟类的感官中以视觉最为发达,听觉次之,嗅觉最为退化。这些特点是与飞行生活密切联系的。视觉为飞翔定向的主要器官。
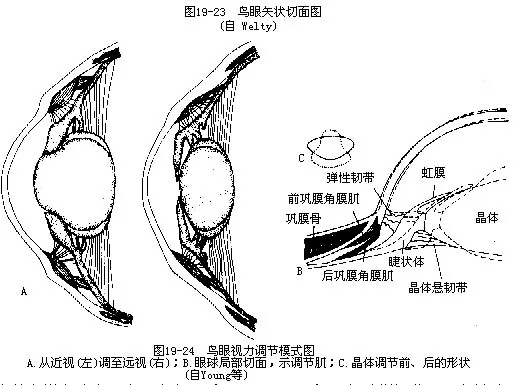
鸟眼的相对大小比所有脊椎动物都大,大多数外观呈扁圆形,为扁平眼(flat eye);鹰类眼球为球状(globular eye),鸮为筒状(tubular eye)。眼球最外壁为坚韧的巩膜,其前壁内着生有一圈覆瓦状排列的环形骨片,称巩膜骨(sclerotic ring),构成眼球壁的坚强支架,使在飞行时不致因强大气流压力而使眼球变形。在后眼房内的视神经背方伸入一个具有色素的、多褶的和富有血管的结构,称为栉膜(pecten)。栉膜的确切功能还不很清楚,它在演化上与爬行类眼内的圆锥乳突(conus papillaris)同源,一般认为有营养视网膜的功能,并可借体积的改变而调节眼球内的压力;也有一些证据指明它可在眼内构成阴影,减少日光造成的目眩(图19—23)。
鸟眼的晶体调节肌肉为横纹肌,此点与除爬行类以外的所有其他脊椎动物不同,对于飞行中迅速聚焦,可能是有利的。眼球的前巩膜角膜肌(anterior sclerocorneal muscle)能改变角膜的屈度,后巩膜角膜肌(posterior sclerocorneal muscle)能改变晶体的屈度(此点类似哺乳类等陆地动物),因而它不仅能改变晶体的形状(以及晶体与角膜间的距离),而且还能改变角膜的屈度,称之为双重调节(图19—24)。鸟类的虹彩呈黄、褐及黑色。虹彩肌也是横纹肌,与哺乳类不同。潜水鸟类在水下时,能借虹彩肌的收缩来压迫晶体前部,协助调焦。
由于鸟类具有这种精巧而迅速的调节机制,使其能在一瞬间把扁平的“远视眼”调整为“近视眼”,鹰类的眼球甚至可被调节成筒状,这是飞翔生活所必不可少的条件。鹰在高空中能察觉田地内的鼠类,并在几秒钟内俯冲抓捕,其视力比人大8倍;燕子在疾飞中能追捕飞虫,这都与具有良好的视力调节分不开的。否则越临近目的物就会越看不清楚。
鸟类听觉器官基本上似爬行类,具有单一的听骨(耳柱骨)和雏形的外耳道。夜间活动的种类,听觉器官尤为发达。

大多数鸟类鼻腔内具有 3个鼻甲(nasal concha),但嗅觉退化。一般认为这也是飞行生活的产物。少数种类(例如兀鹫)嗅觉相当发达,已成为一种嗅觉寻食的定位器官。
(十)生殖鸟类生殖腺的活动,存在着明显的季节性变化。在繁殖期生殖腺的体积可增大几百倍到近千倍。一般认为这也与适应于飞翔生活有关。
1.雄性生殖系统基本上与爬行类相似,具有成对的睾丸和输精管,输精管开口于泄殖腔。鸵鸟和雁鸭类等的泄殖腔腹壁隆起,构成可伸出泄殖腔外的交配器,起着输送精子的作用。在某些鹳形目及鸡形目等鸟类,还残存着交配器的痕迹。这些都可以做为鉴别雌雄性别的标志。但在大多数鸟类,均不具交配器官,借雌雄鸟的泄殖腔口接合而授精。鸟类的精子在泄殖腔和输卵管内存活寿命比哺乳类长,例如将雌家鸭与雄鸭隔离之后,第一周产64%受精卵,第3周为3%,最后一枚受精卵在第17天产出。
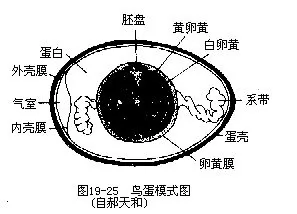
2.雌性生殖系统绝大多数雌鸟仅具单一的(左侧)有功能的卵巢,右侧卵巢退化。但某些鹰类(尤其是雀鹰、鹞和隼)雌鸟有半数个体具有成对的卵巢。一侧卵巢退化,通常认为与产生大型具硬壳的卵有关。成熟卵通过输卵管前端的喇叭口进入输卵管。受精作用发生于输卵管的上端。在受精卵于输卵管内下行的过程中,依次被输卵管壁所分泌的蛋白(albumen)、壳膜(shellmembrane)和卵壳(shell)所包裹。卵在输卵管中移动时,由于管壁肌肉的蠕动而旋转,逐渐被包裹以均匀的蛋白层,两端稠蛋白层随着扭转而成系带(图 19—25)。被系带所悬挂着的卵黄,由于重力关系而使胚盘永远朝上,有利于孵化,这是一种重要的生物学适应。卵壳为碳酸钙(89%~97%)及少数盐类和有机物构成,其表面有数千个小孔,以保证卵在孵化时的气体交换。很多鸟类的卵壳上有各种颜色和花纹,它们是输卵管最下端管壁的色素细胞在产卵前5小时左右所分泌的。卵最后经泄殖腔排出。
幼鸟的输卵管为白线状,产过卵的输卵管虽也萎缩,但上下端的直径不等。这个特点可做为野外工作时鉴定年龄的依据。
光线能刺激家禽提早产蛋以及在秋冬季节产更多的蛋。已知一些野禽如环颈雉、黄腹角雉、麻雀也对光照刺激有反应,而另一些种类则不敏感。增大光照能促进“光敏”鸟类的运动和进食,另外还通过脑下垂体分泌激素刺激卵巢。12~14小时光照对脑下垂体分泌和产卵的刺激最大。有人认为红光的刺激大于白光。有人指出用紫外线(加入微量的可见光)能提高10%~19%产卵率,并增加维生素D的合成。
鸟类具有孵卵、育雏等一系列本能,保证了后代有较高的成活率。
第二节 鸟纲分类
现今已知鸟类约 9 000余种,分为两个亚纲,即古鸟亚纲(Archaeornithes)和今鸟亚纲(Neor-nithes)。
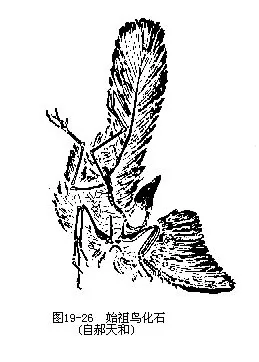
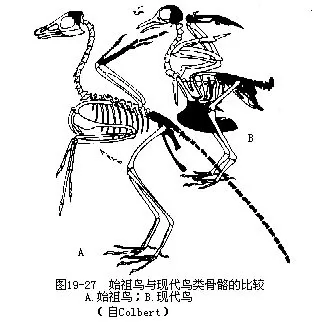
古鸟亚纲以始祖鸟(Archaeopteryx lithographica)为代表,见于距今 1.45亿年前的晚侏罗纪地层中,迄今已报道的化石有7例,均产于德国巴伐利亚省索伦霍芬附近的印板石石灰岩内,即1861(仅为一单根羽毛)、1861(完整骨架,为本种的模式标本)、1877(完整骨架)、1956、1970(系1855年出土)、1973(系1951年出土)和1988年。1984年以来我国发现数具完整的古鸟化石,是德国以外的首次记录。始祖鸟具有爬行类和鸟类的过渡形态。它与鸟类相似的特征是:①具羽毛;②有翼;③骨盘为“开放式”;④后肢具4趾、三前一后。但它又具有类似爬行类的特征,主要有:①具槽生齿;②双凹型椎体;③有18~21枚分离的尾椎骨;④前肢具3枚分离的掌骨,指端具爪;⑤腰带各骨未愈合;⑥胸骨无龙骨突;⑦肋骨无钩状突(图19—26,图19—27)。上述的始祖鸟特征中,除了“羽毛”之外,几乎均可在古爬行类的一些成员中找到,因而曾被称为“有羽毛的爬行动物”,百余年来不时掀起“它究竟是爬行类还是鸟类”的争论, 1985年甚至有人提出“始祖鸟化石标本上的羽毛是伪造的”疑问。经过古生物学家采用多种现代技术手段对标本的鉴定,特别是1988年再次发现具有清晰羽毛印迹的始祖鸟化石标本,才使这一争论告一段落。
1991年印度学者报道了在北美晚三叠纪地层中发现的鸟类化石,订名为原鸟(Protoavis),它具有一些比始祖鸟还原始的、更似恐龙的特征。
对始祖鸟等古鸟化石的研究,关系着鸟类起源于哪一种爬行类的问题。比较流行的观点是鸟类是从近似假鳄类(Pseudosuchia)中鸟龙类(Ornithichia)的槽齿类一支进化而来。 20世纪70年代中期以来,认为始祖鸟更似兽脚恐龙(Theropoda)的腔骨龙(Coelurosauria)的观点曾占上风。不过这一学说近年受到更多的反对,因为不论从化石结构以及生存的地质年代来说,均存在着许多疑问。
今鸟亚纲包括白垩纪以来的一些化石鸟类以及现存鸟类。化石鸟类以黄昏鸟目(Hesperor-nithiformes)和鱼鸟目(Ichthyornithiformes)为代表,它们的骨骼近似现代鸟类但上、下颌具槽生齿。我国近年发现了大量早白垩纪鸟类化石,引起世界瞩目,其中的中国鸟(Sinornis santensis)产于辽宁省辽阳,是世界上已知的最早会飞的鸟;甘肃鸟(Gansus yumenensis)产于甘肃省玉门,是海岸和水鸟的原始类群;华夏鸟(Cathayornis yandica)也在辽阳出土,是除始祖鸟及原鸟以外的最原始的鸟类化石。
现存今鸟亚纲鸟类可归为3个总目,即:
一、平胸总目(Ratitae)
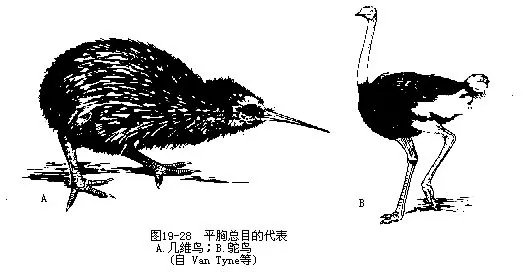
为现存体型最大的鸟类(体重大者达135kg,体高2.5m),适于奔走生活。具有一系列原始特征:翼退化、胸骨不具龙骨突起,不具尾综骨及尾脂腺,羽毛均匀分布(无羽区及裸区之分)、羽枝不具羽小钩(因而不形成羽片),雄鸟具发达的交配器官,足趾适应奔走生活而趋于减少(2~3趾)。分布限在南半球(非洲、美洲和澳洲南部)。
平胸总目的著名代表为鸵鸟(Struthio camelus)或称非洲鸵鸟(图 19—26)。适应于沙漠荒原中生活,一般成小群(40~50只)活动,奔跑迅速。跑时以翅搧动相助,一步可达8m,每小时可跑60km,为快马所不及。食植物、浆果、种子及小动物。雌雄异色,雄鸟背翅色黑。平常所说的“鸵鸟政策”是说“鸵鸟在危急时把头埋在沙堆里”,但后来研究证实,这是一种错误的说法。鸵鸟在繁殖期为一雄多雌,雌鸟把蛋产在一个公共的穴内,每穴可容10~30枚。卵乳白色,重约1300g。孵卵期为6周,白天雌鸟有时孵卵,夜间则由雄鸟担任。
本总目的其他代表尚有美洲鸵鸟(Rhea americana)及鸸鹋(或称澳洲鸵鸟)(Dromaus nova-chollandeae)。此外在新西兰尚有几维鸟(Apteryx oweni),为仅产在此区有限岛屿上的稀有鸟类。体大如鸡,翼与尾均退化,喙长而微弯,鼻孔位于喙的尖端(此点与众不同)。夜出挖取蠕虫等为食,白天钻入地面的洞穴或树根下隐藏。叫声有如尖哨声,并常发出“kiwi…”声,故名几维。产1~ 2枚近白色的卵于洞内,卵的相对大小为鸟类之冠(卵重占体重的1/4)。雄鸟孵卵(图19—28)。
二、企鹅总目(Impennes)
潜水生活的中、大型鸟类,具有一系列适应潜水生活的特征。前肢鳍状,适于划水。具鳞片状羽毛(羽轴短而宽,羽片狭窄),均匀分布于体表。尾短。腿短而移至躯体后方,趾间具蹼,适应游泳生活。在陆上行走时躯体近于直立,左右摇摆。皮下脂肪发达,有利于在寒冷地区及水中保持体温。骨骼沉重而不充气。胸骨具有发达的龙骨突起,这与前肢划水有关。游泳快速,有人称为“水下飞行”。分布限在南半球。
企鹅总目的代表为王企鹅(Aptenodytes patagonicus)(图 19-29)。分布于南极边缘地区,可深入到内陆数百km处集成千百只大群繁殖。繁殖以后可沿海北上至非洲南部。每产一卵,置于冰上。孵卵期约56天。孵卵由雄鸟担任(其他种企鹅为雌雄共同孵卵),此时雄鸟将卵置于脚上,并以下腹部垂下的袋状皮褶将脚面覆盖。企鹅繁殖期配对是在南极的黑夜季节下进行的,待白昼到来前,卵已产出。当雄鸟孵卵时,雌鸟已长途跋涉到海边生活,2个月后再返回。雄鸟在此期间全不进食,靠消耗脂肪维持(最高的绝食记录为4个月)。这一点是哺乳类所不及的。因而企鹅是深入南极冰原内最远的脊椎动物。企鹅虽步行笨拙,但遇警时可将腹部贴地,双翅快速划雪,后肢似活塞般的快蹬,滑行甚速。

由于南极的特殊地理条件(黑夜白昼各半年、一望无际的茫茫雪海和缺乏山林一类的地貌标志)以及企鹅的特点(高度密集繁殖以及失去飞行能力、在陆地易于捕捉),因而近数十年来一直吸引着很多鸟类学家,把它们做为研究鸟类行为的对象。例如,在黑夜条件下,雌鸟、雄鸟及雏鸟是怎样识别的?成鸟依靠什么定位器官得以穿过几百里无外貌特征的水陆返回原巢。有人用10年以上的时间连续到一种企鹅的繁殖地做了973次观察,并将研究对象做了标志。结果发现有 82%的鸟第二年还是原配偶,其中有一对在一起 11年。有关鸟类迁徙定向机制的假说,有的就是根据对企鹅实验的结果加以充实的。
企鹅的主要食物是磷虾、鱼和乌贼等,在极地海域生态系统的能量流转中占重要地位。其所排出的粪便,是极地苔藓、地衣等的主要肥料来源,在土壤形成方面有重要作用。
三、突胸总目(Carinatae)
突胸总目包括现存鸟类的绝大多数,分布遍及全球,总计约35个目,8500种以上。它们共同的特征是:翼发达,善于飞翔,胸骨具龙骨突起。最后4~6枚尾椎骨愈合成一块尾综骨。具充气性骨骼。正羽发达,构成羽片,体表有羽区、裸区之分。雄鸟绝大多数均不具交配器官。
我国所产突胸总目鸟类,计有26目81科。根据其生活方式和结构特征,大致可分为6个生态类群,即游禽、涉禽、猛禽、攀禽、陆禽和鸣禽。现就常见种类略加概述。
(一)?目(Podicipediformes)中等大小的游禽,善于潜水。主要特征为:趾具分离的瓣状蹼。嘴短而钝。羽毛松软如丝,可制上等毛革制品。尾羽几为绒羽构成。在水面以植物茎叶营浮巢。
本目鸟类极善潜水,遇警时能背负幼鸟在水下潜逃,与所有其他游禽不同(潜鸟、秋沙鸭及天鹅可背负幼鸟在水面游逃)。我国常见种类为小?(Podiceps ruficollis)(图19-30),又名水葫芦。体大小似鸽,体羽灰褐色,栖息于水草繁茂的河湖内。杂食性,但以水生动物为主。繁殖期弯折水草,编成浮巢。产近淡黄色或白色钝椭圆形卵, 6~9枚。雌雄均参加孵卵,孵卵期18~ 29天。在孵卵期间,亲鸟离巢时有以绒羽或水草将卵覆盖的习性(雁鸭类也具此习性),有保温及保护的作用。据在我国的观察以及国外报道,这种盖卵习性可能是防御天敌危害的一种适应。我国的乌鸦(特别是小嘴乌鸦)、海鸥以及少数猛禽是危害卵和雏鸟的主要天敌。

(二)鹱形目(Procellariiformes)(信天翁目)大型海洋性鸟类。外形似海鸥,但体型粗壮(大者体长可近1m)。主要特征为:嘴强大具钩,由多数角质片所覆盖。鼻孔呈管状。趾间具蹼。翼长而尖,善于翱翔。产卵于岸边的地上或洞穴中,有时卵下略垫以草叶。卵白色,每产一枚。两性均参加孵卵,孵卵期70~80天。雏鸟尚需哺育42天,为晚成鸟。
我国较常见的种类如短尾信天翁(Diomedea albatrus)。信天翁为漂泊性海鸟,除繁殖期外,几终日翱翔或栖息于海上。环志记载可作8000km的迁飞,并有“环球飞行”的记录(图19—31)。


(三)鹈形目(Pelecaniformes)大型游禽。主要特征有:4趾间具一完整蹼膜(全蹼)。嘴强大具钩,并具发达的喉囊以适应食鱼的习性。我国著名代表有斑嘴鹈鹕(Pelecanus philippensis)和鸬鹚(Phalacrocorax carbo)(图19—32)。
鹈鹕主要分布于热带及温带地区,通体近白色,飞羽暗黑。喉囊特别发达,以此暂存捕获物,并有助于热天散发体温。饲雏期间取食回巢后,将口大张以露出喉囊,诸雏则群集伸头入内竞食。营群巢(树上或地表),产1~4枚白色卵。两性孵卵约6周,孵出后尚需哺育35天。
鸬鹚又称鱼鹰,通体近黑色,杂有花斑。常集成大群捕鱼,数量过高时可对渔业造成危害。集群营巢于树上或岩缝中,产2~4枚淡绿色卵。两性孵卵,孵卵期25天,雏鸟尚需哺育21天。我国自古即驯养鸬鹚捕鱼。但由于此种方法对渔业资源有危害,解放后已不采用。
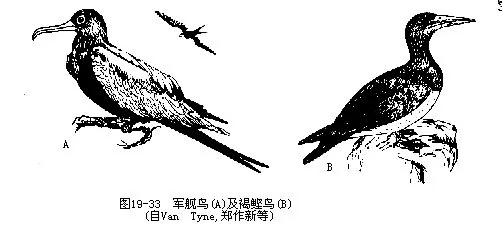
小军舰鸟(Fregata minor)及褐鲣鸟(Sula leucogaster)为我国西沙群岛的著名鸟类,均属热、温带海洋性鸟类。前者是掠夺性鸟类,以其快速敏捷的飞行,于高空夺食鲣鸟、鹈鹕、鸬鹚和海鸥等嘴内所衔鱼类。当它鸟不堪啄击而张嘴时,军舰鸟可在瞬间内把行将落水的鱼类追获啄食。这种习性在鸟类中是罕见的(图 19—33)。
(四)鹳形目(Ciconiiformes)大中型涉禽。栖于水边,涉水生活,嘴、颈及腿均长。胫部裸露。趾细长,4趾在同一平面上(此点与鹤类不同)。雏鸟晚成。
我国常见的有两类,即鹳与鹭。它们外形很相似,但前者中趾爪内侧不具栉状突,颈部不深曲缩成“ S”型(图 19—34)。
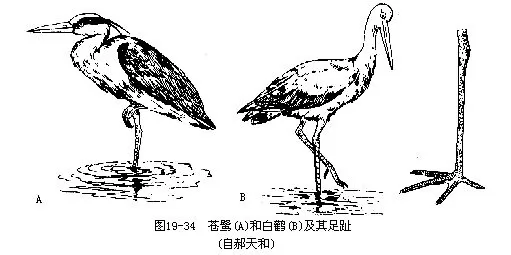
我国常见种类有黑鹳(Ciconia nigra)及东方白鹳(Ciconia boyciana)。在高树或岩崖缝隙内营巢,产3~5枚白卵,孵卵期30~38天。东方白鹳在我国东北繁殖,为世界著名的珍禽,应严加保护。
鹭类常多种集群营巢。其胸腰部侧面长有一种特殊的“粉”,能不断地生长并破碎成粉粒状物,借以清除食鱼时所粘着的污物。中趾的栉状梳即用于梳除粉粒。我国常见种类有大白鹭(Fgretta alba)及苍鹭(Ardea cinerea)。白鹭在我国尚有多种,均属珍贵鸟类,其纯白的矛状羽和蓑羽为贵重的装饰品,许多国家竞相猎取,致使数量显著下降,应大力保护。过去在颐和园内的高树上即有白鹭营巢,巢大而简陋,产3~5枚淡绿色卵。
(五)雁形目(Anseriformes)大中型游禽,是重要经济鸟类。其主要特征为:嘴扁、边缘具有梳状栉板(有滤食功能),嘴端具加厚的“嘴甲”。腿后移,前3趾间具蹼。翼的飞羽上常有发闪光的绿色、紫色或白色的斑块,称为“翼镜”。气管基部具膨大的骨质囊,有助于发声时的共鸣。雄鸟具交配器官。尾脂腺发达。雏鸟孵出后不需哺育即能独立活动(早成鸟)。
雁形目鸟类遍布于全世界,主要在北半球繁殖。多具有季节性的长距离迁徙的习性,其中在我国繁殖、过路及越冬的种类有40余种。通常所说的野鸭、雁及天鹅均属此目(图19—35)。常见代表有:
绿头鸭(Anas platyrhynchos)及斑嘴鸭(Anas poecilorhyncha)为常见的鸭类,均在我国繁殖。家鸭就是从它们驯化而来的。绿头鸭雌雄异色,雄鸟头和颈辉绿色,颈下部有白环,体羽大体灰褐色。雌鸟近棕褐色。斑嘴鸭雌雄均近棕褐色,嘴黑褐色、先端具淡黄色斑块。繁殖于我国河北省北部以北地区,营巢于岸边草丛中,以杂草及绒羽垫衬,产淡绿或淡黄色卵8~12枚。
常见的雁类有鸿雁(Anser cygnoides)及豆雁(Anser fabalis)。体型较鸭类大,雌雄羽色相似,以暗棕色为主。鸿雁嘴比头长,豆雁嘴比头短。鸿雁为我国家鹅的原祖。雁类的一般生活习性似鸭,但陆栖性较强,以植物为主食。在迁徙及越冬期间,有时对冬小麦危害严重,但经济价值大,为重要狩猎对象。
雁形目中体型最大的是天鹅(Cygnus cygnus)。全体洁白,嘴黄具黑斑,游泳时长颈直伸于水面。体姿优美,常被做为文艺作品的主题。此鸟稀少而珍贵,为我国重点保护鸟类之一。
(六)隼形目(Falconiformes)肉食性鸟类,体多大、中型。嘴具利钩以撕裂捕获物。脚强健有力,借锐利的钩爪撕食鸟类、小兽、蛙、蜥蜴和昆早等动物。善疾飞及翱翔,视力敏锐。幼鸟晚成性。白昼活动。雌鸟较雄鸟体大。
我国隼形目鸟类种类及数量均多,羽毛(特别是飞羽及尾羽)有重要经济价值。常见种类有红脚隼(Falco vespertinus),体型大小似鸽,雄鸟背羽灰色,腿脚红色,飞行快捷似燕,又名青燕子。春夏季节来我国繁殖,侵占喜鹊及乌鸦巢产卵,“鹊巢鸠占”的“鸠”即指此鸟。红脚隼在营巢习性上的特点是少见的,但主食害鼠、害虫,是著名的益鸟。鸢(Milvus migrans)也是城乡上空习见的猛禽,体羽褐色,尾羽叉状。鹗(Pandion haliaetus)是一种专以鱼类为食的猛禽,其外趾能后转、各趾下及爪侧均具锋利的鳞状突起,以利于抓捕鱼类。秃鹫(Aegypius monachus)为我国境内的大型猛禽,主要栖息在我国西部及北部的高山上,嗜食动物尸体,头部光秃或仅具绒羽,为此类动物的主要特征(图19—36)。
隼形目鸟类除羽毛具有经济价值外,就食性而论,除少数大型种类捕食野鸭、家禽和家畜等外,绝大多数以鼠类为主食,有益于农田。至于少数种类对于鸟兽的危害方面,多数生态学家认为:①这些猛禽的数量(密度)是有限的,因而至多在局部地区或时期内构成威胁;②它们所抓获的捕获物,大多是老弱病残;从有利于动物的种群更新来看,却具有积极作用。因而隼形目的所有种类均已被列入我国重点保护对象。
隼形目造巢于高树或岩崖缝隙内,产1~6枚近似球形的卵。卵白色或淡绿色,多数带有稀疏不等的褐斑。孵卵由雌鸟担任或由两性负担,孵卵期约35天。幼雏多遍布白色绒毛,尚需哺育40余日始离巢。
猛禽具有吐出“食丸”的习性,即在其栖息地(特别是巢的近旁)休息时,将所吞入的鼠类、鸟类等的不能消化的残团(特别是骨骼及羽、毛)吐出。采集和分析这些食丸,对于查明当地有害啮齿类的种类和数量变动,能提供很有价值的资料。
(七)鸡形目(Galliformes)适应于陆栖步行,与鸠鸽目一起被归入陆禽类。腿脚健壮,具适于掘土挖食的钝爪。上嘴弓形、利于啄食植物种子。嗉囊发达。翼短圆,不善远飞。雌雄大多异色,雄鸟羽色鲜艳,繁殖期间好斗,并有复杂的求偶炫耀。雏鸟早成。
鸡形目为重要的经济鸟类,除肉、羽以外,还有很多种类为著名的观赏鸟,其中有不少是我国特产。我国鸡形目种类十分丰富,而且大多是留鸟,为很多国家及地区所不及,因而合理的狩猎和严格的护、养措施是极为重要的。在我国东北北部的雷鸟(Lagopus lagopus)和榛鸡(Tetrastesbonasia)是北方类型的代表,它们的跗跖部具有羽毛、无距(腿后的刺状物),以及鼻孔被羽,可与其他鸡类相区别。雷鸟适应于苔原地带生活,繁殖期羽毛褐色、冬季变为白色,在雪原中生活,是鸟类保护色的著名事例。绿孔雀(Pavo muticus)、白鹇(Lophura nycthemera)、红腹锦鸡(金鸡)(Chroysolophus pictus)、环颈雉(Phasianus colchicus)、长尾雉(Syrmaticus reevesii)、原鸡(Gallusgallus)等,均为有经济价值并可供观赏的鸟类。褐马鸡(Crossoptilon mantchuricum)为我国特产的稀有鸟类,限产于河北北部及山西北部的局部山林中,是重要的保护对象。原鸡是家鸡的祖先,我国鸟类学家已证明家鸡在我国自古就有驯化,所谓“中国家鸡是从印度引入”的说法并不正确。
鸡形目中尚有一些小型短翼种类,常见的有鹌鹑(Coturnix coturnix)、鹧鸪(Francolinuspintadeanus)、石鸡(Alectoris chukar)等,它们地栖性更强,均为产量甚高的狩猎鸟(图 19-37)。
鸡形目以植物种子为主食(雷鸟、榛鸡等以树的嫩枝叶芽为主食)。繁殖期常为一雄多雌,绝大多数在地面营巢。一般产8~15枚卵,卵色多样。孵卵期20~30天。
(八)鹤形目(Gruiformes)体型大小不等的涉禽。涉禽类的腿、颈、喙多较长,胫部通常裸露无羽,趾不具蹼或微具蹼,4趾不在一平面上(后趾高于前3趾)。鹤形目雏鸟为早成鸟。
本目的著名代表丹顶鹤(Grus japonensis)是世界稀有鸟类之一,在我国东北及内蒙古自治区北部繁殖,是我国重点保护鸟类。鹤的鸣声高吭洪亮与具特殊的发声器有关:它的气管下端盘卷在胸骨附近,并随年龄而逐渐延伸,老鹤可盘成多圈,并穿入胸骨内。营巢于地面,产2卵,两性孵卵32天。雏鸟孵出片刻后即可离巢(此时体重为成鸟的百分之一),这种早熟性是较为少见的。
秧鸡类与骨顶(Fulica atra)也属鹤形目种类,由于数量众多,为狩猎对象。大鸨(Otis tar-da)(俗称地鵏)是能飞翔的鸟类中体重最大的,栖于草原荒地,以奔走为主,在我国东北及内蒙古自治区繁殖。此鸟由于过度猎捕,已很稀少(例如,英国到1838年即已绝迹),是世界濒危物种、我国重点保护鸟类(图19—38)。
(九)鸻形目(Charadriiformes)涉禽,多为中小型鸟类,种类很多,主要分布在北半球。体多为沙土色,奔跑快速。翼尖善飞。雏鸟为早成鸟。由于体色具有隐蔽性、能突然起飞而方向不定,是运动狩猎的主要对象。有的分类系统将鸥形目归入本目。
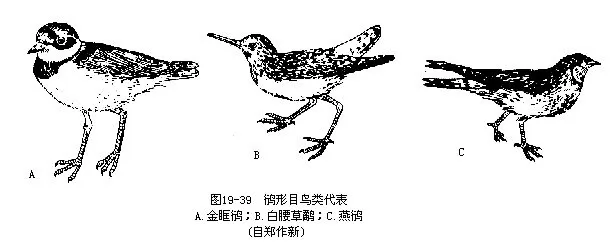
我国常见种类有金眶鸻(Charadrius dubius),为小型鸻类,在河滩上产洋梨形卵4枚,颜色与砂石很相似,虽咫尺之内也难发现。孵卵期约20天。白腰草鹬(Tringa ochropus)为鹬类的常见代表,通常所说“鹬蚌相争,渔人得利”,就是指这类鸟。燕鸻(Glareola maldivarum)嘴短而宽,尾分叉,俗名土燕子,为我国著名的捕食蝗虫的鸟类;据在山东微山湖地区调查,一窝燕鸻一个月内可消灭蝗虫多达16200个(图 19—39)。
(十)鸥形目(Lariformes)海洋性鸟类,与鸻形目亲缘关系密切,但习性近于游禽。常栖息于水边捕食,又似涉禽。体羽大多为银灰色。前3趾间具蹼。翼尖长而善飞翔。雏鸟在形态上为早成鸟,但孵出后留巢待哺,习性似晚成鸟。巢置于地表,产2~4枚洋梨形卵,孵卵期约20天。
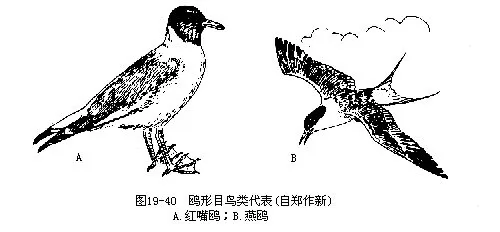
我国常见种类有红嘴鸥(Larus ridibundus)及燕鸥(Sterna hirundo)。后者体似鸽而形似燕,常深入到内陆繁殖(图 19— 40)。鸥与燕鸥常集成大群活动,在渔业区有时可对鱼苗造成危害。但燕鸥(俗称海燕)的多数种类嗜食草地螟等害虫,为非渔业区的益鸟。

(十一)鸽形目(Columbiformes)陆禽。嘴短,基部大多柔软。鼻孔外具有蜡膜(cere)。腿却健壮,4趾位于一个平面上。嗉囊发达,在育雏期能分泌鸽乳喂雏。雏鸟为晚成鸟。本目种类大多为狩猎对象。

我国常见种类有毛腿沙鸡(Syrrhaptes paradoxus),其特征介于鸡与鸽之间。栖于荒漠沙地,在内蒙古自治区繁殖。体沙土色,后趾退化、前3趾并合,覆以毛羽。翼长善飞,常集成千百只大群作不规律地远距离迁飞,寻找水源,1888年我国沙鸡西迁曾达于英国。产卵于地面,卵暗绿色具褐斑,长钝椭圆形。孵卵期28天。此鸟为重要狩猎对象。原鸽(Columba livia)为家鸽的祖先,分布几遍全球,我国仅在南疆西部有分布。山斑鸠(Streptopelia orientalis)和珠颈斑鸠(S.chinensis)均为常见的鸠鸽类。营巢于树上或岩石缝,产2枚白色钝椭圆形卵。两性孵卵,孵卵期14~19天。雏鸟留巢期12~18天,第一周由亲鸟饲以鸽乳(图19-41)。

(十二)鹦形目(Psittaciformes)攀禽。第4趾后转(称对趾型)、嘴坚硬具利钩,上嘴能上抬,均有利于在树上攀缘及掰剥种皮。大多营巢于树洞中。产白色近球形卵,孵卵21天。雏鸟为晚成鸟。
鹦形目为热带鸟类,是著名的观赏鸟。我国云南、广西的极南部以及海南岛所产的绯胸鹦鹉(Psittacula alexandri)可做本目代表。此外,原产澳洲的虎皮鹦鹉(Melopsittaccs undulatus)已广泛作为笼鸟饲养。此目鸟类有多种以谷物为食,数量多时危害严重。善效人言是鹦鹉的著名习性。
(十三)鹃形目(Cuculiformes)攀禽。对趾型。外形略似小鹰,但嘴、爪不具钩。多数分布于欧亚大陆的种类为寄生性繁殖,将卵产于它鸟巢中。雏鸟为晚成鸟。
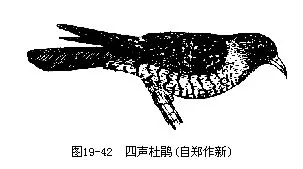
我国常见种类有大杜鹃(Cuculus canorus)和四声杜鹃(C.micropterus)(图 19-42)。前者叫声如“布谷”,故又称布谷鸟;后者叫声加“割麦割谷”。由于杜鹃的规律性迁徙十分显著,每年早春即来,叫声洪亮,彻夜不停,因而自古即已引起人们的注意,并成为各种诗歌、传说的主题。
大杜鹃可在雀形目鸟类(莺科、雀科、画眉科、鸫科、伯劳科、山雀科、鸦科等)约100余种鸟类的巢中产卵,其所产的卵色与义亲的卵很相象。我国最常见的大杜鹃义亲是大苇莺。幼雏孵出早(大约13天),出壳后具有特殊的本能,能将巢内义亲的卵和雏抛出巢外(图19—43),而独受义亲的哺育。杜鹃目为著名益鸟,嗜食松毛虫,这是其他食虫鸟类所比不了的。

(十四)鸮形目(Strigiformes)夜行性猛禽,内部结构近于攀禽。除外形具备猛禽类特征以外,其外趾能后转成对趾型,以利攀缘。两眼大而向前,眼周有放射状细羽构成的“面盘”,有助于夜间分辨声响。听觉为夜间的主要定位器官,耳孔特大,耳孔周缘具皱襞或具耳羽,有利于收集音波。羽片柔软,飞时无声。营巢于树洞中,产白色球形卵1~7枚。雏鸟为晚成鸟。
我国常见种类如长耳鸮(Asio otus)(图 19—44)。脸形似猫,又名“猫头鹰”。鸮类常在深夜发出洪亮而凄厉的叫声,加以外貌不似它鸟美观,因此常被认为是一种不祥之鸟,其实它是著名的食鼠益鸟,食物中的90%以上为鼠类。它这种夜间灭鼠的本领,为隼形目所不及。所有鸮类均已列为我国重点保护鸟类。
(十五)夜鹰目(Caprimulgiformes)夜行性攀禽。前趾基部并合(称并趾型),中爪具栉状缘。羽片柔软,飞时无声。口宽阔、边缘具成排硬毛,适应于飞捕昆虫。体色与枯枝色相似,为白天潜伏时的保护色(图19—45)。不营巢,置 1~ 2枚卵于地表,雏鸟为晚成鸟。
夜鹰嗜食蚊虫(曾在一鸟胃中检出500多只蚊虫),故又名蚊母鸟。近年来屡经证实,夜鹰目种类具有休眠现象,以度过缺食的寒冷季节。我国常见种类为夜鹰(Caprimulgus indicus)。
夜鹰并非文学作品中所说的“夜莺”。后者为小型鸣禽,是与我国著名笼鸟红点颏(红喉歌鸲)(Luscinia calliope)类似的种类。
(十六)雨燕目(Apodiformes)小型攀禽,代表种类雨燕科的后趾向前(称前趾型)。羽多具光泽。雏鸟为晚成鸟。

我国常见种类为楼燕(北京雨燕)(Apus apus)。常集成大群于高空疾飞捕虫,外形颇似家燕,但4趾朝前。金丝燕(Collocalia spp.)繁殖期以唾液腺分泌物营巢,即著名的滋补品“燕窝”。
蜂鸟为世界上最小的鸟类,体重仅1g左右,主要分布于南美洲。以花蜜为食,能在花前似直升飞机般的“悬停”(此时每分钟扇翼达50余次),胸肌相对大小为鸟类之冠(图19—46)。近年发现此鸟在花蜜短缺时期有休眠的事例。
(十七)佛法僧目(Coraciiformes)攀禽。脚为并趾型。种类较多,形体各异。营洞巢,多为白色球形卵,雏鸟为晚成鸟。
本目在我国的常见代表有翠鸟(Alcedoatthis)和戴胜(Upupa epops)。翠鸟嘴形粗大似凿,背羽翠绿色,尾羽短小,以鱼虾为主食。沿岸崖穿凿土穴为巢。戴胜嘴细长而微下弯,以地面蠕虫等小动物为食。头顶具扇状冠羽,俗称花蒲扇。在树洞、建筑物缝隙以及柴堆缝中以草茎编皿状巢,产4~8枚椭圆形污白色卵。孵卵期12~15天。雌鸟在孵卵期间,能自尾脂腺分泌一种特臭的黑棕色液体,对巢、雏有保护作用。
双角犀鸟(Buceros bicornis)为产于我国云南南部的珍贵观赏鸟类。嘴巨大而下弯,上嘴顶部有角质隆起物。在高大树洞中筑巢,孵卵期雌鸟伏于洞内,用雄鸟衔来的泥土混以从胃内呕出的分泌物将洞口封闭,仅留略可伸出嘴尖的洞隙,以接受雄鸟喂食。这是免遭天敌(猴、松鼠及蛇等)伤害的一种适应。孵卵期 28~ 40天。直到雏鸟快出飞时,雌鸟始“破门而出”(图 19-47)。

(十八)?形目(啄木鸟目)(Piciformes)攀禽。脚为对趾型。嘴形似凿,专食树皮下栖居的害虫(如天牛幼虫)。尾羽的尾轴坚硬而富有弹性,在啄木时起着支架的作用。凿洞为巢,产3~5枚白色钝椭圆形卵。孵卵期10~18天,雏鸟为晚成鸟。
啄木鸟为著名的森林益鸟,除其所特有的消灭树皮下面的害虫之外,还可根据其凿木的痕迹做为森林“卫生采伐”的指示剂。因而被称为“森林医生”。我国常见种类有斑啄木鸟(Picoidesmajor)(图 19—48)。

(十九)雀形目(Passeriformes)鸣禽,占现存鸟类的绝大多数(约5千余种)。其主要特征为:鸣管及鸣肌复杂,善于鸣啭。足趾3前一后,后趾与中趾等长(称离趾型)。跗跖后部的鳞片愈合成一块完整的鳞板。大多营巢巧妙,雏鸟为晚成鸟。
雀形目为鸟类中最高等的类群,在鸟类进化的历史上较其他各目出现晚,并处于剧烈的辐射进化阶段,种类繁多(多达64个科)。常见代表有百灵(Melanocorypha mongolica)、家燕(Hirundo rustica)、喜鹊(Pica pica)、秃鼻乌鸦(Corvus frugilegus)、 画眉(Garrulax conorus)、黄眉柳莺(Phylloscopus inornatus)、大山雀(Parus major)、麻雀(Passer montanus)、燕雀(Fringilla montifringilla)等。其中有的是农林益鸟,有的是农业害鸟,还有的因其善于鸣啭和效鸣而为著名的笼鸟(图19—49)。

第三节 鸟类的繁殖、生态及迁徙
一、鸟类的繁殖
鸟类繁殖具有明显的季节性,并有复杂的行为(例如占区、筑巢、孵卵、育雏等),这些都是有利于后代存活的适应。
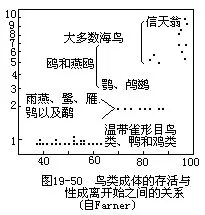
鸟类的性成熟大多在生后一年,多数鸣禽及鸭类通常不足一岁就达到性成熟,少数热带地区食谷鸟类幼鸟经3~5个月即可繁殖。鸥类性成熟需3年以上,鹰类4~5年,信天翁及兀鹰迟至9~12年性成熟。性成熟的早晚一般与鸟类种群的年死亡率相关(图19—50);死亡率愈低的,性成熟愈晚,每窝所繁殖的雏鸟数也少。
大多数鸟类的配偶关系维持到繁殖期终了、雏鸟离巢为止。少数种类为终生配偶,已知的有企鹅、天鹅、雁、鹳、鹤、鹰、鸮、鹦鹉、乌鸦、喜鹊及山雀等。在世界鸟类中,有2%科和4%亚科鸟类是一雄多雌(例如松鸡、环颈雉、蜂鸟及织布鸟);约0.4%科及1%亚科鸟类是一雌多雄(例如三趾鹑及彩鹬);其余大多为一雄一雌。
普通鸟类每年繁殖一窝(brood)。少数如麻雀、文鸟及家燕等,一年可繁殖多窝。在食物丰富、气候适宜的年份,鸟类繁殖的窝数和每窝的卵数均可增多。一些热带地区的食谷鸟类甚至几乎终年繁殖。
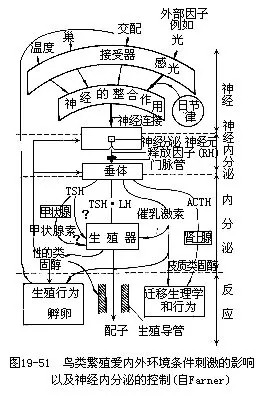
鸟类性腺的发育和繁殖行为的出现,是在外界条件作用下,通过神经内分泌系统的调节加以实现的。每年春季,光照条件的改变以及环境景观的变化等因子,通过鸟类的感官作用于神经系统,影响丘脑下部的睡眠中枢,使鸟类处于兴奋状态。丘脑下部的神经分泌神经元(肽能神经元)向脑下垂体门静脉内分泌释放因子(RF),引起脑下垂体分泌。脑下垂体所分泌的卵泡刺激素(FSH)和黄体生成素(LH)促使卵巢的卵细胞发育并分泌性激素(性的类固醇),使生殖细胞成熟并出现一系列繁殖行为。脑下垂体所分泌的促甲状腺激素(TSH)促使甲状腺分泌甲状腺素,以增进有机体的代谢活动,提高生殖行为的敏感性。脑下垂体所分泌的促肾上腺皮质激素(ACTH)促使肾上腺分泌肾上腺素,提高了有机体对外界刺激的应激能力,有利于完成与繁殖有关的迁徙等行为(图19—51)。鸟类在整个繁殖周期内,雄鸟的求偶炫耀、交配、造巢和孵卵等一系列活动,也都不断地通过感官作用于神经内分泌系统,强化着鸟类性周期的生理活动和行为。日节律(昼夜节律)(circadian rhythm)的体内生物钟,对繁殖周期活动也有影响。
鸟类每年进入繁殖季节以后,随着性腺的发育,出现一系列的繁殖行为,例如向繁殖地区迁徙、占区、求偶炫耀、筑巢、产卵和孵卵以及育雏活动等,待雏鸟离巢之后,亲鸟开始秋季换羽并陆续离开营巢地点,到适宜的地区越冬。现就一些主要内容加以介绍:
(一)占区或领域(territory)鸟类在繁殖期常各自占有一定的领域,不许其他鸟类(尤其是同种鸟类)侵入,称为占区现象。所占有的一块领地称为领域。占区、求偶炫耀(courtship dis-play)和配对(pair formation)是有机地结合在一起的,占区成功的雄鸟也是求偶炫耀的胜利者。
占区的生物学意义主要表现在:①保证营巢鸟类能在距巢址最近的范围内,获得充分的食物供应。所以飞行能力较弱的、食物资源不够丰富和稳定的,以及以昆虫及花蜜为食的鸟类,对领域的保卫最有力;②调节营巢地区内鸟类种群的密度和分布,以能有效地利用自然资源。分布不过分密集,也可减少传染病的散布;③减少其他鸟类对配对、筑巢、交配以及孵卵、育雏等活动的干扰;④对附近参加繁殖的同种鸟类心理活动产生影响,起着社会性的兴奋作用。
领域的大小可从几平方km2(例如鹰、鵰、鹫、和雪鸮)到几万m3(如雷鸟),一些雀形目小鸟的领域约为几百m2。领域大小是可变的,在营巢的适宜地域有限、种群密度相对较高的情况下,领域可被其他鸟类“压缩”或“分隔”而缩小。这在我国华北地区(历经数千年的开发、大量森林被改变为耕地,林区已极度缩小)的某些雀形目鸟类中尤为明显,以致我们可以推断鸟类占区造巢的演化途径或许就是:由于环境条件的改变,适宜巢址有限,以致使营“独巢”的鸟类被迫压缩其领域,而成“松散的群巢”;再进一步压缩,则形成“群巢”。
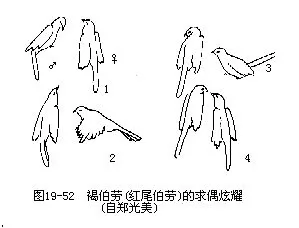
鸟类在占区和营巢过程中,雄鸟常伴以不同程度和不同形式的求偶炫耀(图19—52)。终日在领域内鸣叫(尤以雀形目最为突出)。求偶炫耀和鸣叫都是使繁殖活动得以顺利进行的本能活动,使神经系统和内分泌腺处于积极状态,激发异性的性活动,从而使两性的性器官发育和性行为的发展处于同步(synchronize)。求偶炫耀对于两性的辨认(特别是雌雄同型鸟类)也是十分重要的。有人认为,由于求偶炫耀(例如鸣叫)在鸟类中存在着种的特异性,因而对于亲缘关系较近的不同种鸟类,起着生物学的隔离机制作用,可避免种间杂交。求偶炫耀活动衰退,或被领域附近的新的“入侵者”所超过时,常导致繁殖进程中断。
(二)筑巢(nest-building)绝大多数鸟类均有筑巢行为。低等种类仅在地表凹穴内放入少许草、茎叶或毛;高等种类(雀形目)则以细枝、草茎或毛、羽等编成各式各样精致的鸟巢。鸟巢具有以下功能:①使卵不致滚散,能同时被亲鸟所孵化;②保温;③使孵卵成鸟、卵及雏鸟免遭天敌伤害。鸟类营巢可分为“独巢”和“群巢”两类。大多数鸟类均为独巢或成松散的群巢。群巢在岛屿及人迹罕见的地区最为常见,其中如各种海鸟(企鹅、信天翁、鹈鹕)、鸥类、鹭类、雨燕类及某些鸦科鸟类。鸟类集群营巢的因素是:①适宜营巢的地点有限;②营巢地区的食物比较丰富,可满足成鸟及幼雏的需要;③有利于共同防御天敌。这些因素中,可能“适宜营巢地点不足”是主要原因。随着人类对自然界的大规模开发,适宜巢址的进一步减少,集群营巢的趋势将更加明显。
我国常见的鸟巢,依其结构特点,可分为以下几类(图19—53):
1.地面巢除去某些雀形目鸟类(如百灵、柳莺)也可在地表编织精巧的巢以外,地面巢代表着低等地栖或水栖鸟类(鸵鸟、企鹅以及大部陆禽、游禽、涉禽)的巢式。巢的结构简陋,卵色与环境极相似,孵卵鸟类也具同样的保护色。
2.水面浮巢某些游禽及涉禽能将水草弯折并编成厚盘状,可随水面升降。
3.洞巢产卵于树洞或其他裂隙内,一些猛禽、攀禽及少数雀形目鸟类属此。洞穴的位置、结构与鸟类的生活习性有密切关系。其中较低等的种类都不再附加巢材,产白色卵(反映了原始森林鸟类似爬行类的卵)。雀形目洞巢种类则于洞内置似复杂的巢材,卵色也多样,反映出是一种后生的特化现象。


4.编织巢以树枝、草茎或毛、羽等编织的巢。低等种类(如鸠鸽自、鹭类、猛禽)的巢型简陋。雀形目鸟类则能编成各种型式(皿状、球状、瓶状)的精致鸟巢。我国以造巢著名的鸟类有织布鸟(Ploceus philippinus)和缝叶莺(Orthotomus sutorius)。前者以嘴将植物纤维如织布般地穿梭编织成瓶状巢;后者以植物纤维贯穿大形树叶的侧缘,而缝合成悬于树梢的兜状巢。
随着人类的出现,有不少鸟类(特别是洞巢鸟类)已转入在建筑物上营巢。对于这些与人类接触较密切的鸟类,要注意研究其益害。
(三)产卵(egg-laging)与孵卵(Incubation)卵产于巢内并加以孵化。卵的形状、颜色和数目(以及卵壳的显微结构、蛋白电泳特征)在同一类群间常常是类似的,从而也可以反映出不同类群之间的亲缘关系,可做为研究分类学的依据。
每种鸟类在巢内所产的满窝卵数目称为窝卵数(cluth)。窝卵数在同种鸟类是稳定的,一般说来,对卵和雏的保护愈完善、成活率愈高的,窝卵数愈少。就同一种鸟而言,热带的比温带的产卵少;食物丰盛年份的产卵数多。所以窝卵数是自然选择所赋予的、能养育出最大限度的后代数目。此外,窝卵数也与孵卵亲鸟腹部的孵卵斑所能掩盖的数目有关。
鸟类中存在着定数产卵(determinate layer)与不定数产卵(indeterminate layer)两种类型,前者在每一繁殖周期内只产固定数目的窝卵数,如有遗失亦不补产,例如鸠鸽、鲱鸥、环颈雉、喜鹊和家燕等。不定数产卵者,在未达到其满窝卵的窝卵数以前,遇有卵遗失即补产一枚,排卵活动始终处于兴奋状态,直至产满其固有的窝卵数为止。已知一些企鹅、鸵鸟、鸭类、鸡类、一些啄木鸟以及一些雀形目鸟类(例如家麻雀)均有此特性;饲养卵用家禽(鸡、鹌鹑、鸭、鹅及火鸡等)就是利用了鸟类的这种特性。
孵卵大多为雌鸟担任(例如伯劳、鸭及鸡类等),也有的为雌雄轮流孵卵(如黑卷尾、鸽、鹤及鹳等),少数种类为雄鸟孵卵(如鸸鹋、三趾鹑等)。雄鸟担任孵卵者,其羽色暗褐或似雌鸟。除少数种类(例如企鹅、鸬鹚、鸭及鹅)之外,参与孵卵的亲鸟腹部均具有孵卵斑。孵卵斑有单个的(例如很多雀形目鸟类、猛禽、鸽及?)、两个侧位的(例如海雀及鸻形目鸟类)以及一个中央和两个侧位的(例如鸥与鸡类)。孵卵斑的大小与窝卵数多少之间没有什么相关。
已测鸟类孵卵时的卵温为34.4℃~35.4℃左右。在孵卵早期,卵外温度高于卵内温度;至胚胎发育晚期,卵内温度略高于卵外温度。
每种鸟类的孵卵期通常是稳定的,一般大型鸟类的孵卵期较长(如鹰类29~55天、信天翁63~81天、家鸽18天、家鸡21天、家鸭28天、鹅31天),小型鸟类孵卵期短(例如一般雀形目小鸟为10~15天)。
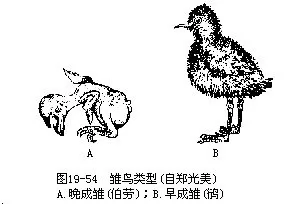
(四)育雏(Parental care)胚胎完成发育后,雏鸟即借嘴尖部临时着生的角质突起——“卵齿”将壳啄破而出。鸟类的雏鸟分为早成雏(precocial)和晚成雏(al-tricial)。早成雏于孵出时即已充分发育,被有密绒羽,眼已张开,腿脚有力,待绒羽干后,即可随亲鸟觅食。大多数地栖鸟类和游禽属此。晚成雏出壳时尚未充分发育,体表光裸或微具稀疏绒羽,眼不能睁开。需由亲鸟衔虫饲喂(从半个月到8个月不等),继续在巢内完成后期发育,才能逐渐独立生活(图19—54)。雀形目和攀禽、猛禽以及一部分游禽(体躯大而凶猛的种类,如鹈鹕、信天翁)属此。可以看出,雏鸟早成性或晚成性,是长期自然选择的产物;凡筑巢隐蔽而安全,或亲鸟凶猛足可卫雏的鸟类,其雏鸟多为晚成雏。早成雏是地栖种类提高成活率的一种适应性。尽管如此,早成雏的卵与雏的死亡率都比晚成鸟高得多,因而产卵数目也多。
晚成雏的发育,一般表现为“S”型生长曲线,即从早期的器官形成和快速生长期过渡到物质积累和中速生长期,至晚期的物质消耗大于积累生长期。在雏鸟发育早期,尚缺乏有效的体温调节机制,需靠亲鸟伏巢来维持雏鸟的体温。随着雏鸟内部器官的发育,产热和神经调节机制的完善以及羽衣(体温覆盖层)的出现,而转变为恒温。对我国多种鸟类雏鸟体温发育测定的结果,发现在羽衣的羽鞘破裂、形成羽片的当日,常是恒温出现的转折。例如褐伯劳(Lanius cristatus lu-cionensis)的羽鞘破裂期为孵出后10日龄,对其体温的测定(半导体温度计测泄殖腔温度)结果为:

很多种晚成雏(例如企鹅、鹈鹕、信天翁、雨燕、鹦鹉、翠鸟及食蜂鸟等)在离巢前的体重超过成鸟,为脂肪积累所致。这种适应有助于雏鸟渡过由于阴雨等因素所造成的食物短缺,并为离巢前的飞羽、肌肉等的生长提供较充分的能量。由于雏鸟在离巢前活动频繁、能量消耗巨大,常见有体重显著下降现象。阴雨是造成雏鸟大量死亡的一个重要因素。
晚成雏鸟类在育雏期的食量很大,而且以昆虫为主食,此期大多数的种类有益于农林。
二、鸟类的迁徙
迁徙(migration)并不是鸟类所专有的本能活动。某些无脊椎动物(如东亚飞蝗)、某些鱼类、爬行类(如海龟)和哺乳类(如蝙蝠、鲸、海豹、鹿类)也有季节性的长距离更换住处的现象。其中海龟与鲸的迁徙距离可从数百km到上千km。但是作为整个分类类群来说,鸟类的迁徙是最普遍和最引人注目的,因而多年来一直成为动物学研究的一个重要领域。
鸟类的迁徙是对改变着的环境条件的一种积极的适应本能;是每年在繁殖区与越冬区之间的周期性的迁居。这种迁飞的特点是定期、定向而且多集成大群。鸟类的迁徙大多发生在南北半球之间,少数在东西方向之间。
根据鸟类迁徙活动的特点,可把鸟类分为留鸟(resident)和候鸟(migrant或transient)。留鸟终年留居在出生地(繁殖区),不发生迁徙,例如麻雀、喜鹊等。候鸟则在春、秋两季,沿着固定的路线,往来于繁殖区与越冬区域之间,我国常见的很多鸟类就属于候鸟。其中夏季飞来繁殖,冬季南去的鸟类称夏候鸟(summer resident),如家燕、杜鹃;冬季飞来越冬、春季北去繁殖的鸟类称冬候鸟(winter resident),如某些野鸭、大雁。夏季在我国某地以北繁殖,冬季在我国某地以南越冬,仅在春秋季节规律性地从我国某地路过的鸟类称旅鸟或过路鸟(如极北柳莺等)。
严格的说,现今所说的留鸟,有不少种类秋冬季节具有漂泊或游荡的性质,以获得适宜的食物供应,有人称这种鸟为漂鸟(wanderer)。
(一)迁徙的原因 引起鸟类迁徙的原因很复杂,至今尚无肯定的结论。大多数鸟类学者认为,迁徙主要是对冬季不良食物条件的一种适应,以寻求较丰富的食物供应,尤其以昆虫为食的鸟类最为明显。此外,有人认为北半球夏季的长日照(昼长夜短)有利于亲鸟以更多的时间捕捉昆虫喂养雏鸟。这2种意见可以相辅相成,但是还不能解释有关迁徙方面所涉及的各种复杂事实。
有人从地球历史来推测鸟类迁徙的起源问题。在新生代第四纪(约10万年前)曾发生多次冰川运动,自北半球向南侵袭。冰川所到之处,气候剧变,冰雪遍地,不利于鸟类生存。冰川周期性的侵袭和退却,使鸟类形成了定期性往返的生物遗传本能。从这种认识出发,提出2种互相对立的假说:①现今的繁殖区是候鸟的故乡,冰川到来时迫使它们向南退却,但遗传的保守性促使这些鸟类于冰川退缩后重返故乡,如此往返不断而形成迁徙本能;②现今的越冬区是候鸟的故乡,由于大量繁殖,迫使它们扩展分布到冰川退却后的土地上去,但遗传保守性促使这些鸟类每年仍返回越冬区(故乡)。
冰川说并不能解释有些鸟类为什么不迁徙。而且有人指出冰川期(第四纪更新世)仅占整个鸟类历史的百分之一,因而它对鸟类遗传性的影响终究是有限的。所以也还不能排除在冰川期以前鸟类即已存在着迁徙的事实。
(二)迁徙的诱因 曾有不少假说来解释鸟类迁徙的诱发因素,例如认为光照、食物、气候以及植被外貌的改变,都可以引起迁徙活动。现今较大量的实验证明,光照条件的改变,可以通过视觉、神经系统而作用于间脑下部的睡眠中枢,引起鸟类处于兴奋状态。光刺激还增强了脑下垂体的活动、促进性腺发育和影响甲状腺分泌,增强机体的物质代谢,进一步提高对外界刺激的敏感性,从而引起迁徙。
我们认为,迁徙是多种条件刺激所引起的连锁性反射活动。其中物种历史所形成的遗传性是迁徙的“内因”,外界刺激为引起迁徙的“条件”。
(三)迁徙的定向(Orientation) 迁徙的最显著特点是每一物种均有其较固定的繁殖区和越冬区,它们之间的距离从数百km到千余km不等。而且实验证明,很多鸟类(例如家燕、企鹅)次年春天可返回原巢繁殖。即使是用飞机将迁徙鸟类运至远离迁徙路线的地区内,释放数天之后仍可返回原栖地。因而就存在着“鸟类究竟依靠什么来定向”的问题。根据野外观察、环志、雷达探测、月夜望远镜监视以及各种各样的室内实验,曾提出不少假说,但均处于探索阶段,尚未获得肯定结论。目前比较流行的看法有:
1.训练和记忆 认为鸟类具有一种固有的、由遗传所决定的方向感(an innate sense of di-rection),这种方向感,随着幼鸟跟随亲鸟迁徙的过程中,不断地加强对迁徙路线的记忆。
2.视觉定向(visual orientation)依靠居留及迁徙途径的地形、景观(例如山脉、海岸、河流以及荒漠等)作为响导,并不断从老鸟领会传统的迁徙路线。实验表明,视觉定向对于鸟类短距离的归巢,可能不是主要的。例如在鸽眼上装以额接触镜,然后于距巢15km~130km处放飞,大部分仍能按时归巢,可见一定存在着视觉以外的定向机制。
3.天体导航(celestial navigation)很多实验表明,鸟类能利用太阳和星辰的位置定向。星辰定向对于夜间迁徙的鸟类尤为重要。
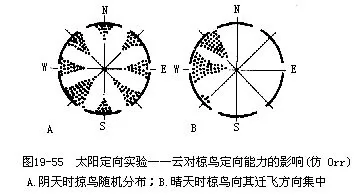
关于太阳定位的实验,最著名的是克拉默(Kramer,1957,1961)用椋鸟作的研究。把具有迁徙习性的椋鸟,放在四面有窗的笼内,以激素处理使其进入迁徙状态,则可见椋鸟朝着一定的方向(即其迁徙方向)扇翼(图19—55),而且扇翼行为在阴天不出现。如果用镜子代换太阳的方位,则扇翼方向可按人所预定的方向变更。把企鹅移至远离巢区的茫茫雪原内释放之后,则于晴天沿直线走向原居住地;在阴天则乱走;一俟天晴,又立即寻获正确方位。在太阳定位方面,有人认为鸟类根据太阳的方位角(sun azimuth)来确定方向。有人认为鸟类是借当地中午时太阳的顶空高度与其所记忆的原居住地的情况加以比较,来测定所在的纬度。有人认为以日落的方向定标,再根据星辰、风向等加以校正。这些均是尚待充分验证的假说。
星辰定向是由索尔(Sauer,1957)首次在圆形笼内对欧洲苇莺进行实验得出的,证明它们能根据夜空中星辰的位置定向。此后又做了大量实验研究,而且用改变人造星辰位置的方法,也可以像上述的太阳定向的实验一样,使鸟类按预定的方向改变其“迁飞”方向。
把太阳及星辰作为航行的指针时,由于地球的自转,有机体必须具有一个内部的“生物钟”(biological clock)借以不断调整太阳及星辰与其迁徙轴之间的角度。近年有些实验证明,当以人工光照改变白天—黑夜周期,可使鸟类产生定向错误(“生物钟”被重新调整了),当把该鸟再置于真正的日光照射条件下时,仍按错误的时间和方向迁飞。这方面的工作还有待于继续深入研究。
4.磁定向(magnetic orientation)认为鸟类能借地磁感应来确定迁徙方向。这个学说最早是用将鸟笼放在四周为铁壁的室内,能使鸟类丧失方向的定向性而提出来的(Fromme,1961)。以后相继有大量的实验,特别是德国鸟类学家(Merkel,Wiltschko,1965;Wiltschko et al,1971—1981)作了引人注目的研究:他们以八角形鸟笼饲养一些迁飞的鸟类(欧莺),鸟笼外置以能产生强力人工磁场的线圈(helmholta coils),能选择性地改变人工磁场的方向。实验证实,即使在鸟类能看到满布星辰的夜空情况下,它们也随着人工磁场方向的改变而变更着“迁徙”的方向。他们认为夜间迁徙鸟类的方向选择,主要是靠对磁场的感应,而迁徙方向的保持则与星辰位置有关;也就是说,星辰用于校准地磁罗盘的定向,星辰定向是基于磁定向的信息。
有人(Southern,1972)在鸥的幼鸟头上装以陶瓷磁铁,发现其定向机能被干扰。实验用雷达干扰带有磁片的家鸽,能使之丧失“归巢”定位。还有人用微波辐射(microwave radiation)照射小鸡,以实验寻找一种减少迁徙鸟类对飞机威胁的方法。这是因为多年来在不同高度(2000m~7000m)的上空,均发生过集群迁徙的鸟类(例如雁鸭类)与飞机相撞而造成机毁人亡的事故。实验初步证实,鸟类的羽毛能对微波辐射起反应。这些还有待进一步研究。
除了上述的依靠地形、景观、天体、磁场等定向之外,目前还有大量资料(包括人造地球卫星所摄制的照片)证明,鸟类在一定的地理条件下,能依靠气象条件(主要是季节性的风)来选择迁徙方向,并借助风力进行迁徙。所有这些事实及假说,还均处于探索阶段,对于彻底揭示鸟类迁徙与定向之谜,还有待于深入研究。
(四)研究迁徙的意义 对迁徙机制及其迁飞途径的研究,不论在理论上和实践上都具有重要意义。在理论上能够揭示迁徙本能的形成及其发展过程,为生物进化论以及有机体与环境之间的复杂关系提供更为深入的资料。在实践上除了为有效地利用和控制经济鸟类,为改造自然区系提供理论基础外,还为仿生学提供了广阔的研究领域。现今所设计制造的定向导航系统,尽管已日益精确,但从某种意义上说,还远不如生物定向系统。
第四节 鸟类与人类的关系
本节试图以鸟类为例,通过讨论来阐明动物在人类生活中的重要性、怎样判断益害以及保护和合理利用野生动物资源方面所涉及的复杂性。
家养动物有史以来给人类生活所带来的巨大利益及其在人类生活中的重要性,已是不言而喻的,迄今世界各国仍一直在寻求和培育新的驯养品种,以满足日益增长的社会需求。以鸟类而论,除了家鸡、家鸭、鹅、火鸡、珠鸡、鹌鹑等是早已从野生原祖驯化而来的以外,近年来大量引入我国并广泛饲养作为肉、蛋、皮革及羽用的鸵鸟、孔雀、环颈雉以及“美国鹧鸪”[实为原产欧、亚大陆的石鸡(Alectoris gracea)]等,都是近几十年培育出的新家禽品种。探求将有巨大经济价值和驯养繁殖前景的野生动物变为家养,具有广阔的前景,也是保护和利用野生动物资源的一种途径,需要有大量动物学家来从事这项研究。至于家禽品种的进一步培育和饲养增殖,则是遗传学家和农学家的主要任务。这里仅就野生鸟类与人类的关系作些介绍。
绝大多数鸟类(以及野生动物)是有益于人类的,它们是维护人类的生存环境、以及生态系统稳定性的重要因素。近年来,生物多样性的保护问题已成为全球关注的热点之一,1992年联合国环境与发展大会上通过了《生物多样性公约》,我国是最早缔约国之一,承担了保护我国生物多样性的义务,其中当然包括野生动物。这实质上是人们对野生动物“利”与“害”认识方面的一个根本性的转变。从历史发展看,人类对事物的认识是随着科学的发展而不断革新和深化的。早期人们考虑野生动物的益和害时,视野比较狭窄,往往只是看到与人类的直接利害,例如食、用价值高不高,是吃害虫还是吃庄稼,是否传布疾病等。随着研究的深入才发现,不仅在回答上述问题时要涉及非常复杂的因素,需要进行大量深入的科学研究,而且当把一个物种作为生态系统中的成员来加以考虑时,就会知道我们对所面临的问题了解得太少,有些只是皮毛,要有许多工作去做。从生态系统的稳定性和生物多样性保护这一基本原则出发,对野生动物、特别是目前尚缺全面认识的绝大多数野生动物,要妥善地加以保护,在此基础上进行科学的、合理的永续利用。对于局部地区和时间内造成危害的动物,要在科学指导下进行适当的控制。“引入”或“消灭”一个物种,要采取极为审慎的态度。
保护的目的在于利用,在使生态系统保持相对稳定的、健康的良性物质循环的基础上,要合理的、最大限度的利用动物资源。取用那些有经济价值的、每年通过繁殖而增长的种群中的剩余部分。否则让其自生自灭也是一种浪费。鸟类在动物类群中是益处极大、害处极小的一个类群,除了所提供的生态效益和经济效益之外,它在科学和社会文明的发展上有重要贡献。生物进化理论以及许多生物学和生态学的理论,都是首先从鸟类学研究中揭示并进而在其他类群中得到验证的。鸟类在城市园林中的点缀及其在文学、艺术创作方面的贡献,更是众所周知的。因而“爱鸟”和“观鸟”早已成为先进国家的一种广泛的群众运动。
鸟类与人的直接利害关系主要有:
一、鸟类的捕食作用
(一)对捕食作用的估价大多数鸟类能捕食农林害虫,即使是主食植物性食物的鸟类,在繁殖期间也以富含营养及水分的昆虫(特别是鳞翅目幼虫)来饲喂雏鸟,在抑制害虫种群数量的增长上有相当的作用。猛禽是啮齿动物的天敌,许多小型猛禽也主食昆虫,因而在控制鼠害和虫害、清除动物的尸体和降低动物流行病的传布等方面,都有重要作用。鸟类的种类和数量众多,分布于多种生态环境内,特别是飞行生活的习性使之能追随集群移动的蝗虫、鼠类等的机动性捕食能力,是其他捕食动物类群所不可比拟的。所以从总体上看,特别是从整个生态系统中鸟类的作用来考虑,要对食虫鸟类和猛禽予以全面的保护,已是世界的共识。
然而要判断每一种鸟类在具体地区和时间内的捕食作用,也就是它究竟有多大益处,却是难以回答的,必须进行大量的科学研究,而不能简单地以某鸟一天的食量来估算出全年能消灭多少害虫或害鼠,甚至再折算出相当于保护了多少粮食。以食虫鸟类而论,实涉及:①食虫鸟类所吃的虫子中有多少是害虫,有多少是捕食性昆虫(益虫),通过捕食之后对二者之间的关系有无影响;②在特定地区内食虫鸟类的种群数量及其主要猎物(害虫)的数量动态,特别是在鸟类捕食前后的猎物种群密度。事实上判断鸟类捕食效果(益处大小)的唯一标准是“通过鸟类的捕食作用,其主要猎物的种群密度是否已被抑制在不致为害的水平”。以昆虫的繁殖潜力而论,如果天敌不能使其种群密度降低到90%以上,所残留的个体会通过繁殖而迅速地恢复到原有水平。
就是由于上述问题的难度,迄今达到这一深度的研究成果不多。不过就已有的材料可以认为,鸟类在害虫的密度较低时,有较明显的捕食作用,它能阻滞或防止害虫的大发生或延长大发生的间隔期。例如对美国卷叶蛾的数量动态研究发现,在正常年景大约有20%~65%幼虫被鸟捕食,而在害虫大发生时仅被捕食3.5%~7.0%。我国浙江马尾松人工林内的大山雀对松毛虫的捕食,在一般年景为4.71%~22.19%,而在松毛虫大暴发的年份仅为0.22%,降低20倍。这主要是由于食虫鸟类种群数量的增长远远低于害虫的增长。尽管已知某些鸟类在猎物丰盛的年份可以借提高繁殖力以扩大种群,但与其猎物的增长相比,是微不足道的。有人计算在美国某林区的卷叶蛾大发生年份,其种群数量增大8000倍,而对此反应最强的栗胸林莺(Dendroica cas-tanea)仅增加12倍。当然,在自然界捕食某种害虫的不止一种鸟类。由于食虫鸟类的种类多、分布广,其对害虫的抑制作用,特别是在维持正常年景下的生态系统的稳定方面,是相当重要的。同时也应认识到,在林业经营中企图单纯靠鸟类去控制虫害是不现实的。这特别是由于大多数人工林以幼林居多、林型单一,其所吸引和栖居的鸟类本来就十分稀少。所以在森林害虫的防治工作中应提倡综合防治的策略,即:发展低成本的高效、无残毒化学杀虫剂,利用多种天敌生物(病毒病原体、真菌、捕食及寄生性昆虫、食虫鸟类等)以及林型的合理配制。任何单一的防治方法均有其局限性。
(二)食虫鸟类的保护与利用 保护食虫鸟类的根本原则是保护和改善它的栖息环境,控制带有残毒的化学杀虫剂的使用以及禁止乱捕滥猎。这是一件长期的任务,要广泛开展宣传教育工作,提高全社会的认识。我国自1988年颁布《中华人民共和国野生动物保护法》和1991年以来每年开展“爱鸟周”活动以来,已经收到了相当显著的效果。
在园林地区悬挂人工巢箱来招引食虫鸟类,为那些在洞穴内筑巢的种类提供更多的巢址,是国内外早已广泛采用的方法,特别在缺乏树洞的幼林内有比较明显的效果。但是对悬挂人工巢箱招引食虫鸟类的措施也要适度,并不是悬挂巢箱愈多招引来的鸟类就愈多。这首先是由于食虫鸟类中只有少数种类是在洞穴内筑巢并喜欢选用人工巢箱的;其次是食物资源或环境载力的制约,在有限的条件下不可能允许食虫鸟类种群数量无限增多。迄今国内外的研究均证实,麻雀是小型鸟类中的霸主,在有麻雀分布的城市园林内的人工巢箱中,90%以上被麻雀所侵占。所以在城市园林地区悬挂人工巢箱主要更应着眼于通过这些活动对青少年以及社会风尚所带来的积极影响。“十年树木、百年树人”的思想对于野生动物的保护工作是很恰当的。一些宣传媒介所谓的某城市公园通过“招鸟工程”使鸟类在几年内“增加了17倍”的报道,就是对上述常识缺乏了解的一种浮誇风。
我国近年广为兴起的“驯鸟放飞捉虫”活动,也是一种劳民伤财的、对群众性爱鸟热情的误导。首先,所驯养的灰喜鹊、红嘴蓝鹊等并不是典型食虫鸟,而是杂食性鸟类;其次,这种“养兵千日、用在一时”的作法既耗费人力、物力,又易造成在饲养中的死亡;而且在现场放飞时,放飞人员所能达到的地点极为有限,有限环境所能提供的食物也是有限的。“哨声一响就回笼”的原因就在于笼内食物优于自然界。所以这是一种驯鸟杂技表演而不是“生物防治”。即使是对于真正的食虫鸟类,唯一可行的保护和利用途径也应是在自然界内予以保护,使其自食其力地繁衍生息,而不必耗费大量财力及人力去强当“保姆”。把爱鸟活动从笼内转变为大自然,需要广泛深入的科学普及工作以及对传统的某些社会意识进行变革。
(三)鸟类捕食对植物散布的影响 许多鸟类是花粉的传播及植物授粉者,例如蜂鸟、花蜜鸟、太阳鸟、啄花鸟、锈眼鸟等。以植物种子或果实为食的鸟类,都会有一些未经消化的种子随粪便排出,这些经过鸟类消化道并与粪便一起排出的种子更易于萌发,会随着鸟类的飞移而广为散布。已知一些海洋岛屿上的植物就是经由鸟类扩散到的。星鸦、松鸦及某些啄木鸟有在秋季贮藏植物种子的习性,可将数以百计的针叶树球果或栗树种子贮藏到数km以外的树洞内,有人认为这是历史上欧、美栗林扩展的主要原因。
二、狩猎鸟类
狩猎鸟类主要包括一些鸡形目、雁形目、鸠鸽目、鸻形目以及一些秧鸡、骨顶等。它们都是种群数量增长较快的、有季节性集群的以及肉、羽等经济价值高的鸟类。在对其繁殖力及种群数量动态进行充分研究的基础上,合理狩猎会带来巨大的经济收益。
运动或休闲狩猎在许多发达国家甚为流行,在规定的狩场和猎期内定量狩猎是一种娱乐,国家发放狩猎证以及其他的服务性收入,每年获利数以亿美元计。为了适应这种需要,有专门的研究机构对猎物的生态学和种群数量动态进行长期的研究,人工饲养、繁殖大批猎物(例如环颈雉、灰山鹑等),定期释放到野外供狩猎之用。所以这既是一种保护野生动物资源下尽量满足人类文化生活需要的措施,也是将谷物等转化为高蛋白肉质品的一种经营方式。
三、鸟害
鸟类所造成的危害常是局部的,因时、因地以及人们的认识程度和具体需求而异。最明显的是农业鸟害,例如雁、鹦鹉、雉、鸠鸽以及雀形目中的鸦科、雀科、文鸟科的许多种类都嗜食谷物或啄食秧苗,其中最著名的就是麻雀。这要在权衡得失的基础上,选择适宜的方法加以控制。“人、鸟争食”的矛盾在生产力水平以及生活水平比较低的情况下十分尖锐,随着农作技术水平的提高以及社会需求的变化,在认识上会有所改变的。变害为益也是可能的。前文提到的放养雉类似供狩猎的做法就是一例。
“鸟撞”(bird strike)是飞机航行中与大群迁徙鸟类相撞而引发的事故,通常多发生在航机起、落或作低空飞行的情况下。自从60年代以来由涡轮喷气发动机取代了螺旋桨推进器之后,由涡轮机进气口将飞鸟吸进而引起空难的事故日益增加。因此在机场的选址(特别是沿海机场的选址)时要了解该地迁徙鸟类的种类、出现季节和飞迁方向、飞行高度等。机场建成之后也应对鸟类的活动规律进行全天、全年的监测。要通过对机场附近生态环境的改造以及发展一些物理、化学及生物的综合技术进行驱鸟工作。
鸟类可以携带一些细菌、真菌和寄生虫等,有些可在家禽、家畜或人类之间传布。世界上曾有十几个国家流行过鸟热病,死亡率高达1/3以上。迄今已知与鸟类有关的传染病有20余种。因而开展鸟类疾病与寄生生物的研究,查明它们之间的传播途径以及与人类健康的关系,对于鸟类的保护和人类的健康都是十分迫切的。
复习题
1.总结鸟类主要特征以及与爬行类相似的要点。
2.总结鸟类适应飞翔生活方式,在各个器官系统上的结构特点。
3.鸟类进步性特征表现在哪些方面?
4.鸟类的3个总目在分类特征上有哪些主要区别?
5.总结鸟类的各种生态类群由于适应不同的环境和生活方式,在形态结构上有哪些趋同性特征?
6.始祖鸟化石的发现有何意义?它具备哪些特征?
7.什么叫迁徙?举例说明留鸟和候鸟。
8.鸟类繁殖行为有哪些特征?试述其生物学意义。
9.简述鸟类与人类的关系。
本文标题:第十九章 鸟纲(Aves)
手机页面:http://m.dljs.net/dlsk/dongwuxue/59414.html
本文地址:http://www.dljs.net/dlsk/dongwuxue/59414.html
很赞哦!(21757)
上一篇: 第五人格紫金皮肤兑换码是什么
站长推荐








友情链接
- 弹弹堂大冒险技能训练隐身 弹弹堂大冒险技能优先升级
- 热血传奇:能爆出麻痹戒指的三种怪物,一种绝版,还有两种需要挖
- 皮肤图鉴 | SHU033周仓
- 冒险王3OL辅助相关手游新闻
- 三国志战略版巨戟时出现在哪里
- 《烟雨江湖》大昭寺势力商店有什么
- 俄乌战争打出一个大国难题,中国已成普京王牌
- 烟雨江湖云纹玄晶怎么获得 烟雨江湖云纹玄晶获得攻略
- 时空猎人0.1折版 5.1.992 安卓版
- 疯狂骑士团试练塔最新出装 试练塔怎么打攻略
- 王者荣耀8月14号皮肤碎片商店更新:曹操幽灵船长值得兑换?
- 航海王燃烧意志女帝攻略大全 女帝加点及玩法指南
- 2019lol终极皮肤 lol赏金终极皮肤值不值得买?好
- 英雄联盟策士统领斯维因玩法介绍及皮肤推荐
- 绿色征途手游折扣平台 低价充值渠道
- 方舟生存进化皮肤代码有哪些-皮肤代码大全2023
- 摩尔庄园手游精灵怎么召唤 摩尔庄园精灵召唤方法
- 摩尔庄园手游澡堂位置介绍
- 动物学上学期知识点总结
- 烟雨江湖追寻神秘人攻略
- 火线精英 11月24日23:00更新维护公告
- 赛尔号12月5日活动汇总
- 五代十国第一明君 立志30年平天下 他若不死何来弱宋强辽
- 赛尔号玄武超进化活动的时间是什么
- 绷带下的魅力 版本第一打野阿木木解析
- 阴阳师双鸩阵容(阴阳师PVE阵容)
- 三国志战略版:个性系统最佳选择?同盟计策频出?pick出你的最爱
- 梦幻西游三维版安卓12月18日公测上线 多重豪礼相赠挚友
- 赛尔号雷伊的倒影怎么打
- 约战精灵再临
- 胡莱三国2ipad版
- 天龙八部自动打怪脚本 v4.0 绿色免费版
- 传阿汤哥追求米兰达可儿:离公开不远了
- 热血江湖里师傅叫什么
- 摩尔庄园精灵宝宝怎么抓
- 赛尔号一路有你:第四期-给小火猴洗特性特性和雇佣怎么用赛尔号启航手游3
- 胡莱三国真孟获技能搭配全解析
- 冒险王3OL下载手机最新版本 2023冒险王3OL手游极速安装教程
- 新晋百万级纯电轿跑,仰望U7四电机四驱,最高车速270km/h
- 赛尔号SPT boss系列(14)被遗忘的SPT还是被包装成SPT的活动精灵?元素之灵布兰登